Revision
of the Castlemainian isograptid (graptolite) biostratigraphy in western
Newfoundland
Jörg Maletz1,
Charles E. Mitchell1, Michael J. Melchin 2 and Tammy Dunlavey1
1Department
of Geology, State University of New York at Buffalo, Buffalo, New York 14260-3050,
U.S.A. E–mail: jorgm@buffalo.edu
2 Department of Earth Sciences, St. Francis Xavier University, P.O. Box 5000, Antigonish, NS B2G.2W5, Canada.
Key words: Graptolites. Isograptus. Biostratigraphy. Middle Ordovician. Western Newfoundland.
Introduction
Since Cooper’s (1973) revision of the genus Isograptus, species of this group have been used as the prime faunal elements to differentiate the Castlemainian (mid Arenig) time interval in outer shelf and slope settings around the world. The use of isograptids, especially of the FAD of Isograptus victoriae lunatus, to define a GSSP at the base of the Middle Ordovician Series has been discussed more recently. Problems with the definition of a GSSP based on conodonts and shelly faunas in the classical Whiterock sections of Laurentia (Finney and Ethington, 2000, Albanesi and Carrera, 2001), and their correlation with the graptolite record (Mitchell, 2001), has resulted in the search for alternative sections.
The Cow Head Group of western Newfoundland contains a unique succession in which conodont and trilobite–bearing limestones are interbedded with isograptid–bearing shales. This succession also has produced the best–documented record of isograptids and pseudisograptids of Castlemainian age known world–wide (Williams and Stevens, 1988). Despite this fact, Williams and Stevens (1988) encountered difficulties in their effort to differentiate the individual subspecies of the I. victoriae lineage and, therefore, did not use the Isograptus victoriae maximus and Isograptus victoriae maximodivergens biozones. They treated the corresponding interval as a broadly defined I. victoriae maximus Biozone. We agree with the problems of the differentiation of I. v. maximus and I. v. maximodivergens and treat them as a single form, I. v. maximodivergens, but differentiate the biostratigraphic units based on the occurrence of associated pseudisograptids.
In addition, Williams and Stevens (1988) considered the I. v. victoriae Biozone to be confined to the interval above Bed 12 at The Ledge (CHN), Jim’s Cove (CHS) and St. Paul’s Inlet (SPI), whereas Bed 12 is within the zone at Martin Point North (MPN) and Martin Point South (MPS). These features, combined with the need to refine the relationship between the graptolite succession and the contemporaneous conodonts faunas, has led us to the re–investigate these faunas of the Cow Head Group (Figure 1). Together with Dr. Svend Stouge, Geological Survey of Denmark, Mitchell and Melchin made coordinated collections from these units in August, 2002. We are presently engaged also in a general reassessment of isograptid systematics based on geometric morphometrics techniques, including the previous and new collections from western Newfoundland. We report here preliminary results of our reassessment of the graptolite biostratigraphy of the Castlemainian interval in the Cow Head Group.
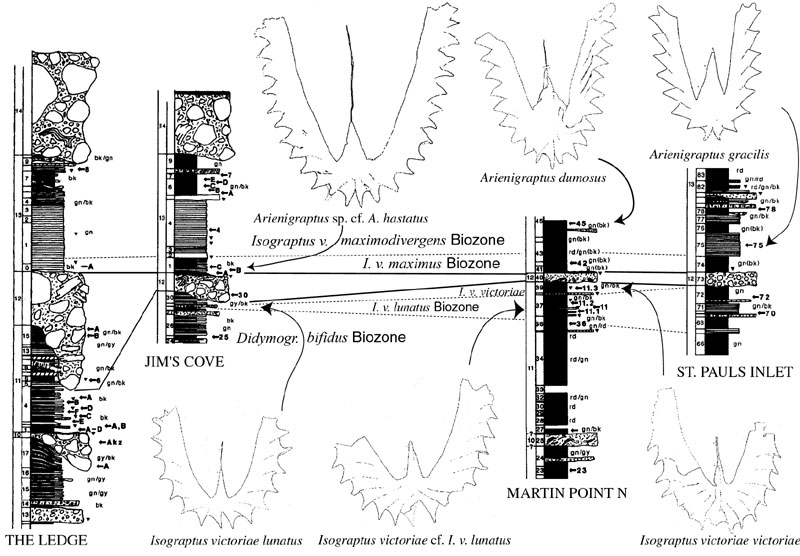
Figure 1. Biostratigraphic correlation of important Cow Head Group sections and indicative faunal elements. Lithostratigraphy from Williams and Stevens (1988).
Cow Head Peninsula (CHN, CHS sections)
These are particularly critical sections for determining the first appearance of the Whiterockian shelly succession. Ross and James (1987) described elements of the earliest Whiterockian Orthidiella Biozone fauna from clasts within the conglomerates of beds 12 to 14 in this region. They inferred from this pattern and from other data elsewhere that the first appearance of the Orthidiella Biozone fauna was likely to be coeval with the age of the shale that encloses the oldest of these beds: Bed 12. Our data support Williams and Stevens’ (1988) identification of the Didymograptellus bifidus Biozone as the youngest fauna below Bed 12 in the CHN section (‘The Ledge’). The parted limestone interval directly above Bed 12 bears a sparse and not very well preserved graptolite fauna. Material described as I. v. victoriae by Williams & Stevens (1988) from this interval in The Ledge section (CHN13/0A; Figures 2A,B,D-E) are mostly juveniles, but are consistently larger in their general dimensions with a considerably longer supradorsal part of the sicula than is typical of I. v. victoriae specimens from Australasia or those found in the Cow Head succession below Bed 12 (e.g., at Martin Point, see discussion below). Among some of the specimens (e.g., Figure 2B) the stipes are relatively slender or even taper distally. In this respect they resemble P. caduceus imitatus. The remaining larger specimens (e.g., Figure 2A,E) exhibit stipes that widening distally as in I. v. maximodivergens. The specimens, however, are too poorly preserved for a precise identification.
Williams and Stevens (1988) also reported Arienigraptus hastatus from this same collection from just above Bed 12 at The Ledge. Among the Australasian collections (which admittedly are few and mostly are isolated sites not well located in continuous stratigraphic sections), Arienigraptus hastatus is restricted to the Ca3, whereas Arienigraptus gracilis and Arienigraptus dumosus first appear in the Ca4 (VandenBerg and Cooper, 1991; Cooper and Ni, 1986). Parisograptus caduceus imitatus also makes its first appearance within the Ca3 – I. v. maximus Biozone. Although less definitive than we would prefer, taken together the fauna from the basal part of Bed 13 (?I. v. maximodivergens, ?Parisograptus caduceus imitatus, and Arienigraptus sp. cf. A. hastatus) appears to us most likely to represent the Ca3 and not the Ca2 – I. v. victoriae Biozone as previously thought. We discuss this issue further, below.
At Jim’s Cove (CHS), Bed 12 erodes less deeply into the underlying succession and specimens of I. v. lunatus were found by Williams and Stevens (1988). We were unable to recollect definitive material of I. v. lunatus from this site but agree with their interpretation of the previously collected material. As at The Ledge, the faunas above Bed 12 in the CHS section contain A. hastatus together with material that we refer to I. v. maximodivergens rather than I. v. victoriae. Thus, we question whether the I. v. victoriae Biozone is present above Bed 12 in the Cow Head Peninsula sections.
Martin Point (MPN and MPS sections)
The Martin Point sections represent a more distal facies in which the Bed 12 conglomerate is relatively thin. Both sections show a good graptolite record below Bed 12. Bed 11/37 at MPN bears an isograptid fauna with large I. victoriae cf. I. v. lunatus specimens (Figure 1). These are similar to typical I. v. lunatus, but are conspicuously larger. Comparable material was figured by Cooper (1973), but included in the intraspecific variation of I. v. lunatus. Williams and Stevens (1988) also included this material in I. v. lunatus. True I. v. victoriae occur in Bed 11/39, still below conglomerate Bed 12. Comparable faunas also can be found in MPS (Williams and Stevens 1988). These specimens agree in every respect with the Australasian material that Cooper (1973) included within this taxon and with specimens collected by Mitchell and Maletz at stop 20 in the Vinini Creek section, Roberts Mountains, Nevada (Finney et al., 1995). There are no graptolites in the red and green shales immediately overlying the Bed 12 conglomerate. Further correlation to Martin Point and St. Paul’s Inlet are mostly based on the lithostratigraphic succession following the practice and usage of James and Stevens (1986) and Williams and Stevens (1988). Younger Castlemainian faunas are known only from higher up in the MPS section, where I. v. maximodivergens Biozone faunas with I. v. maximodivergens, Arienigraptus dumosus, and A. gracilis occur in the upper part of bed 58.
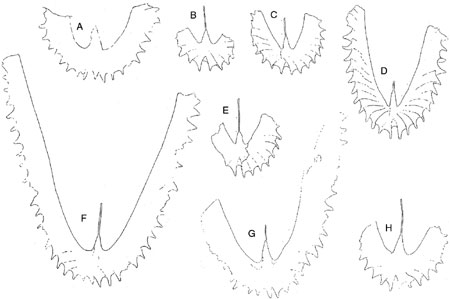
Figure 2. ?I. v. maximodivergens. A, GSC 81387 (Williams and Stevens 1988, Figure 57w). G, GSC 81394 (Williams and Stevens 1988, Figure 57x). H, GSC 81382 (Williams and Stevens 1988, Figure 57u). I. v. victoriae. B, MPN 11, 30cm below base of bed 12, coll. C.E. Mitchell. C, on slab with GSC 81731 (Williams and Stevens, 1988, pl. 19, Figure 4). D, GSC 81728 (Williams and Stevens, 1988, Figure 57s). E, MPN 11, 30cm below base of bed 12, coll. C.E. Mitchell. Specimens show consistently shorter supradorsal length of sicula and webbing between sicula and th11. ?P. caduceus imitatus. F, GSC 81395a (Williams and Stevens, 1988, Figure 57y). Magnification: x5.
St. Paul’s Inlet (SPI section)
The section does not bear useful graptolite faunas directly below Bed 12 and, thus, the correlation of these strata with the I. v. lunatus Biozone by Williams and Stevens (1988) is conjectural. Above the conglomerate an interval with green shale also did not deliver any useful graptolites. The succeeding interval of parted limestone (Bed 13/75), which Williams and Stevens (1988) referred to the Ca2, bears a rich Ca4 – I. v. maximodivergens Biozone fauna. Among the new material we have collected are many mature, well–preserved, and fully typical specimens of I. v. maximodivergens, Parisograptus caduceus imitatus, A. gracilis, and A. dumosus. Williams and Stevens (1988) described isolated juvenile and proximal end fragments of an isograptid that they obtained from SPI75C as I. v. victoriae. Considering the new material we have collected from these same beds and the relatively incomplete suite of characters presented by the isolated specimens, we expect that they are more likely to be specimens of P. caduceus imitatus than I. v. victoriae.
Western Brook Pond (WBN and WBS sections)
The WBN section does not bear any graptolite faunas of the early Castlemainian. The record starts well above Bed 12 with the I. v. maximodivergens Biozone in Bed 13/39. The WBS section, however, has a more complete record. A modest I. v. lunatus Biozone assemblage is present in Bed 11/28 several meters below Bed 12 (Williams and Stevens 1988), which we have also recollected. The presence of the I. v. victoriae Biozone in the green and red shale interval (Bed 11/29) immediately below Bed 12 cannot not proven as these rocks again do not yield any graptolites. They do, however, contain a K–bentonite bed of about 3 cm in thickness. The strata directly above the Bed 12 conglomerate again are green and red shales that are also barren of graptolites. The first indicative faunas above the conglomerate come from Bed 13/33, and this and many succeeding strata bear abundant material of I. v. maximodivergens, I. c. imitatus, A. dumosus, and A. gracilis, indicating a Ca4 age.
Conclusions
1) The I. v. lunatus Biozone is present below the conspicuous Bed 12 conglomerate in the Cow Head Group, as are faunas of the I. v. victoriae Biozone in distal sections. The basal contact of Bed 12, however, is highly diachronous and cuts down into the D. bifidus Biozone at the more up–slope sections of the Cow Head Peninsula (e.g., at The Ledge section, CHN, Williams and Stevens 1988: Figure 4). The only unquestionable occurrences of the I. v. victoriae Biozone in the Cow Head Group are restricted to strata below Bed 12.
2) The oldest well–dated strata known above the Bed 12 unconformity most likely belong to the I. v. maximus Biozone (e.g., the basal part of Bed 13 at CHN and CHS). Slightly higher strata in Bed 13 at St. Paul’s Inlet (SPI75) certainly belong to the I. v. maximodivergens Biozone. In the other localities of the Cow Head Group, the graptolite succession resumes only above an unfossiliferous interval with faunas of the I. v. maximodivergens Biozone.
3) The species succession in the Cow Head Group (especially that of the Isograptus, Arienigraptus and Parisograptus species) appears to match that of the Australasian succession, however, additional data from other regions are needed to confirm this. Again, typical I. v. victoriae from the Vinini Formation at stop 20 (Finney et al., 1995) are not associated with other isograptids or pseudisograptids. Finney and Ethington (1992), however, indicated the presence of I. v. victoriae in typical Yapeenian to Darriwilian assemblages, but did not support the identification with any illustrations. The morphometrics work that we are undertaking should also add significantly to this discussion.
4) The relations described here indicate that Bed 12 is mid–Castlemainian in age but its precise location in the Ca2–Ca3 interval cannot be determined from the graptolite evidence. If we accept that the oldest occurrence of Orthidiella Biozone fossils in the Cow Head conglomerates are approximately coeval with the actual time of origin of this fauna in the shelf environs up slope, then, this basal Whiterockian assemblage is also mid–Castlemainian in age not Ca2 as previously inferred from these rocks. Conodonts from the Cow Head limestones that are currently under study by Stouge may provide a test of this conjecture.
5) The early Castlemainian (Ca1–2) strata below Bed 12 contain a succession of conodont– and trilobite–bearing limestones that together with the graptolite fauna may permit precise correlation of this interval with rocks from a range of basinal and outer shelf facies in many regions around the globe. Thus, the Cow Head succession holds real promise for resolving some of the remaining uncertainties associated with the definition of the base of the Middle Ordovician Series.
References
Albanesi, G.L. and Carrera, M.G. 2001. Niquivil section of central Precordillera, Argentina, proposed as the global stratotype for the base of the Middle Ordovician Series. Geological Society of America, Abstracts with Programs, 33(6): A446–A447.
Cooper, R.A. 1973. Taxonomy and evolution of Isograptus Moberg in Australasia. Palaeontology, 16: 45–115.
Cooper, R.A. 1979. Ordovician geology and graptolite faunas of the Aorangi Mine area, north west Nelson, New Zealand. New Zealand Geological Survey Paleontological Bulletin, 47: 1–127.
Cooper, R.A. and Ni, Y. 1986. Taxonomy, phylogeny, and variability of Pseudisograptus Beavis. Palaeontology, 29: 313–363, pls. 24–27.
Finney, S.C. and Ethington, R.L. 1992. Whiterockian graptolites and conodonts from the Vinini Formation, Nevada: Biostratigraphic implications, pp. 153–169. In: Webby, B.D. and Laurie, J.R. (eds.) Global Perspectives on Ordovician Geology. A.A. Balkema, Rotterdam, Brookfield.
Finney, S.C. and Ethington, R.L. 2000. Global Ordovician Series boundaries and global event horizons, Monitor Range and Roberts Mountains, Nevada. In: Legeson, D.R., Peters, S.G. and Lahern, M.M. (eds.) Great Basin and Sierra Nevada. Boulder, Colorado, Geological Society of America, Field Guide, 2: 301–318.
Finney, S.C., Berry, W.B.N. and Murphy, M.A. 1995. Post–Meeting Trip – Great Basin Graptolites. In: Cooper, J.D. (ed.) Ordovician of the Great Basin: Fieldtrip Guidebook and volume for the Seventh International Symposium on the Ordovician System. Fullerton, California, The Pacific Section Society for Sedimentary Geology (SEPM), 78: 133–151.
James, N.P. and Stevens, R.K. 1986. Stratigraphy and correlation of the Cambro–Ordovician Cow Head Group, western Newfoundland. Geological Survey of Canada Bulletin, 366: 1–143.
Mitchell, C.E. 2001. New graptolite collections from the proposed Whiterock Global Stratotype section: Implications for correlation of the Narrow Section and an alternative proposal for the location of the Middle Ordovician GSSP level. Internet site of Ordovician Stratigraphy Discussion Group (http://seis.natsci.csulb.edu/ordstrat2/default.htm).
Ross, R.J.Jr. and James, N. P. 1987. Brachiopod biostratigraphy of the Middle Ordovician Cow Head and Table Head groups, western Newfoundland. Canadian Journal of Earth Sciences, 24: 70–95.
VandenBerg, A.H.M. and Cooper, R.A. 1992. The Ordovician graptolite sequence of Australasia. Alcheringa, 16: 33–85.
Williams, S.H. and Stevens, R.K. 1988. Early Ordovician (Arenig) graptolites of the Cow Head Group, western Newfoundland, Canada. Palaeontographica Canadiana, 5: 1–167.
Received: February 15, 2003
Accepted: June 15, 2003