The post–persculptus and pre–ascensus graptolite fauna in Scania, south–western Sweden: Ordovician or Silurian?
Tatyana
N. Koren’1, Per Ahlberg2 and Arne T. Nielsen3
1A.P. Karpinskii All-Russian
Geological Research Institute, Sredny 74, 199106, St.-Peterburg, Russia.
E–mail: koren@vsegei.sp.ru
2 Department of Geology,
University of Lund, Sölvegatan, 13, SE-223 62 Lund, Sweden.
3 Geological Museum, Öster Volgade 5-7, DK-1350, Copenhagen, Denmark.
Key words: Graptolites. Zonation. Ordovician/Silurian boundary. Scania. Sweden.
Introduction
This presentation aims at a continuation of the discussion initiated by Melchin et al. (1998) concerning global correlation of the lowermost part of the Silurian based on graptolites. The existed definition of the Ordovician–Silurian boundary at the base of the acuminatus Zone in Dob’s Linn was distinguished by the joint appearance of A. ascensus Davies and P. acuminatus (Nicholson) sensu lato at the stratotype section (Cocks, 1985). Melchin and Williams (2000) showed that A. ascensus appears approximately 1.5 m below P. acuminatus s.s. at Dob’s Linn and that the separate ascensus and acuminatus zones should be recognized at the GSSP.
Previous studies
Since the time of Tullberg and Moberg the Brachiopod Shale or Upper Dicellograptus Shale (Ashgill) and Rastrites Shale (Llandovery) were used as regional stratigraphic units in Southern Sweden. Historically, the uppermost part of the Brachiopod Shale above the Dalmanitina mucronata Zone was assigned by Tullberg in 1883 to the "Zone with C. scalaris" and considered as the uppermost part of the Ordovician. In 1910 Moberg included the "Zone with C. scalaris" into the basal part of the Rastrites Shale. Based on graptolites from the Kullatorp core this unit was correlated with the G. persculptus Zone of Britain (Waern , 1948). The persculptus and acuminatus zones were recognized in the lower part of the Rastrites Shale (Series) in the Ostra Tommarp area (Grahn, 1978) and Lingegård core Nº 27, in western Scania (Nilsson, 1979), while the ascensus Zone fauna was reported from the Lovisfred core (Nilsson, 1984). Recently, the dark shales and mudstones of the Rastrites Shale were assigned to the Kallholn Formation (Bergström, S.M. and Bergström J., 1996).
Biostratigraphy
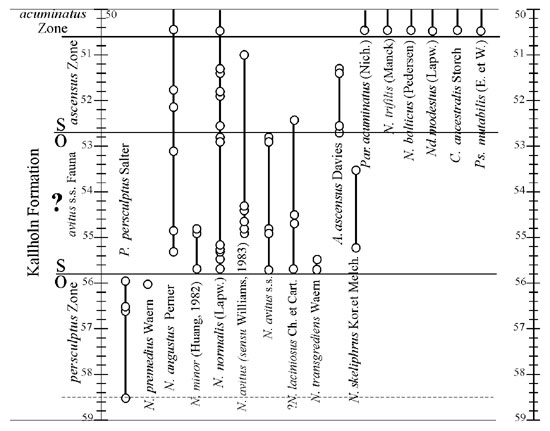
Based on preliminary data the persculptus and acuminatus s.l. biozones were recognized at the lower part of the Kallholn Formation in the Röstanga core (Bergström et al., 1999). Within the black shale succession the O/S boundary was placed at the base of the combined P. acuminatus Zone (incl. A. ascensus Zone) at the depth of 52.70 m, where Akidograptus ascensus first appears (Figure 1). At present, following thorough sampling of the Ordovician–Silurian part in the Röstanga core at intervals as close as 10 cm a succession of four graptolite faunas within the depth of 58 m to 50 m can be recognized (Figure 1).
Figure 1. Range chart of diplograptids in the O/S boundary beds, Röstanga core.
The oldest P. persculptus fauna (58.50 m – 56.10 m.) appears just above the Dalmanitina –bearing beds, which form the uppermost part of the Lindegård Mudstone. It includes numerous and well preserved Persculptograptus persculptus (Salter), Normalograptus angustus (Perner) and single specimens of N. premedius (Waern). This is followed by the N. avitus (Davies) s.s. fauna, lacking akidograptids (55.90 m – 52.70 m). It is differentiated by the first appearances of ?Normalograptus laciniosus (Churkin and Carter), N. avitus s.s., N. minor (Huang) and N. normalis (Lapworth). The fauna mostly consists of the morphologically monotonous and numerically abundant normalograptids, such as N. angustus, N. avitus s.s., N. normalis s.s. and a wider form of N. normalis transient to N. medius (Törnquist), N. skeliphrus Koren and Melchin and N. transgrediens (Waern).The less numerous N. minor and N. avitus (sensu Williams, 1983) are ornamented with a long virgella, bifurcating at different distance from the sicula aperture. Numerical abundance of graptolites increases to the upper part of the interval. The N. avitus s.s. fauna is followed by the A. ascensus association (52.70 m – 50.50 m), distinguished by the FAD of A. ascensus (Davies), the key but not dominant species within this interval. The assemblage is dominated by N. angustus and N. normalis, some rare specimens of N. avitus (sensu Williams, 1983) and N. laciniosus are also found. The P. acuminatus graptolite fauna (50.50 m – 46.70 m) is recognized by the FAD of numerically abundant Parakidograptus acuminatus. This level coincides with the sharp increase in the diplograptid diversity which is seen in the appearance of Normalograptus balticus (Pedersen), N. trifilis (Manck), Cystograptus ancestralis (Storch) and Neodiplograptus modestus (Lapworth). A morphologically characteristic diplograptid species, similar to Talastograptus tuberculatus (Nicholson) does not appear earlier than in the upper zonal interval.
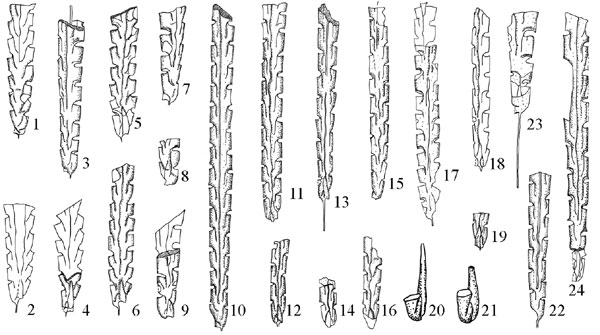
Figure 2. The Persculptograptus persculptus Fauna. 1–9, P. persculptus; 10–22, N. angustus; 23, N. cf. N. avitus s.s., 24, N. premedius. 20–21, x10, all others, x5.
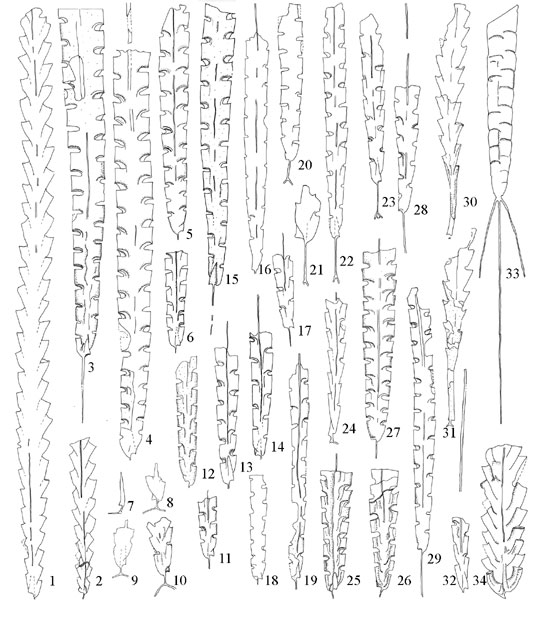
Figure 3. 1–23, the Normalograptus avitus Fauna: 1, 2, N. laciniosus; 3, 4, N. normalis (a wider form); 5, 6, N. normalis s.s., 7–10, N. avitus (sensu Williams, 1983); 11–14, N. angustus; 15, N. transgrediens; 16, 17, N. avitus s.s.; 18, 19, N. skeliphrus; 20–23, N. minor. 24–29 the Akidograptus ascensus Fauna: 24, A. ascensus; 25–27, N. normalis (a wider form); 28, N. avitus s.s.; 29, N. angustus. 30–34, the Parakidograptus acuminatus Fauna: 30, 31, P. acuminatus; 32, N. avitus s.s.; 33, N. trifilis; 34, C. ancestralis. 6, 7, 8, 23, x10, all others, x5.
The same succession of four graptolite faunas was recognized within the O/S boundary interval in the Lönstorp–1 core, Scania (Koren and Nilsen, in prep.) and in the Billegrav–1 core, South Bornholm (Koren and Bjerreskov, 1997). The post–persculptus and pre–ascensus normalograptids of very similar taxonomic composition are present in both cores.
Discussion
The post–persculptus diplograptid fauna lacking akidograptids was recently discovered in Arctic Canada, Algerian Sahara, South Kazakhstan, Kurama Range of Uzbekistan (Melchin et al., 1998; Koren and Melchin, 2000; among the others). In some of those regions the absence of akidograptids in those associations was attributed either to the changes in depositional environments, or to varying ecological parameters. These factors may not be valid in Baltoscandia where there is no evidence of the lithological changes or of a gap in the black shale succession at the O/S boundary. The occurrence of the N. avitus s.s. fauna, which includes neither P. persculptus, no A. ascensus leads to some difficulties in precise definition of the O/S boundary in Baltoscandian succession. Two suggestions can be made. The first, akidograptids make their appearance higher in the study area, and the N. avitus s.s. fauna can be considered to be contemporaneous with the lower A. ascensus fauna elsewhere. In this case the corresponding interval can be included into the ascensus Zone as it was earlier suggested (Nilsson, 1984). This will lead to the difficulties to choose a proper indicator for the O/S boundary. The second, the N. avitus fauna is older than the lower ascensus fauna and the corresponding interval has to be assigned to the Late Ordovician persculptus Zone. In this case, the occurrences of P. persculptus will be limited to the lower part of the zone. Of these two suggestions the only second one can be considered. Then, the lower boundary of the Silurian in Baltoscandia will coincide with the FAD of A. ascensus s.l. and with the base of the ascensus Zone as it is redefined now in Dob’s Linn (Melchin and Williams, 2000).
Acknowledgements
Profs. S. Bergström and K. Larsson are thanked for the kind invitation to study Silurian graptolites of Scania. T.N.K.’s work on the Röstanga and Lönstorp–1 core sampling was funded through the Department of Geology of the Lund University and Geological Museum of the University of Copenhagen. Russian Foundation for Fundamental Science, grant N 02 05–64 775 is to be also acknowledged.
References
Bergström, S.M. and Bergström J. 1996. The Ordovician–Silurian boundary successions in Östergötland and Wästergötland, S. Sweden. GFF, 118: 25–42.
Bergström, S.M., Huff, W.D., Koren, T.N, Larsson, K., Ahlberg, P., and. Kolata, D.R. 1999. The 1997 core drilling through Ordovician and Silurian strata at Röstanga, S. Sweden: Preliminary stratigraphic assessment and regional comparison. GFF, 121: 127–135.
Cocks L.R.M. The Ordovician–Silurian boundary. Episodes, 8: 98–100.
Grahn, Y. 1978. Chitinozoan stratigraphy and palaeoecology at the Ordovician–Silurian boundary in Scåne, southernmost Sweden. Sveriges Geologiska Undersökning, C744: 1–16.
Koren’, T. and Bjerreskov, M. 1977. Early Llandovery monograptids from Bornholm and the southern Urals: taxonomy and evolution. Bulletin of the Geological Society of Denmark, 44: 1–43.
Koren’, T.N. and Melchin, M.J. 2000. Lowermost Silurian graptolites from the Kurama Range, Eastern Uzbekistan. Journal of Paleontology, 74 (6): 1093–1113.
Melchin, M.J, Koren’, T.N. and Williams S.H. 1998. Global correlation of the lower part of the earliest Silurian Akidograptus ascensus–Parakidograptus acuminatus Zone. Temas Geologico–Mineros ITGE, Madrid, 23: 107–108.
Melchin, M.J. and Williams, S.H., 2000. A restudy of the akidograptine graptolites from Dob’s Linn and a proposed redefined zonation of the Silurian stratotype. Geological Society of Australia, Abstract Nº 61. Palaeontology Down–Under: 63.
Nilsson, R. 1979. A boring through the Ordovician–Silurian boundary in Western Scania, south Sweden. Sveriges Geologiska Undersökning, C733: 1–58.
Nilsson, R. 1984. The Didymograptus hirundo and A. ascensus Zones of the Lovisfred core, NW Scania, South Sweden. Geologiska Föreningens I Stockhom Forhandlingar, 105: 261–267.
Waern, B. 1948. The Silurian strata of the Kullatorp core. Bulletin of the Geological Institution of the University of Uppsala, 32: 433–473.
Received: February 15, 2003
Accepted: June 15, 2003