Organic–walled
microfossils at the Silurian/Devonian boundary stratotype (Klonk near
Suchomasty, Barrandian area, Czech Republic)
Oldrich Fatka1, Rainer Brocke2 and Volker Wilde2
1Charles
University, Institute of Geology and Palaeontology, Albertov 6, 128 43 Prague
2, Czech Republic. E–mail: fatka@mail.natur.cuni.cz
2 Research Institute Senckenberg, Palaeobotany, Senckenberganlage 25, D-60325 Frankfurt/Main, Germany.
Key words: Organic–walled microfossils. Stratotype. Silurian–Devonian boundary. Barrandian area. Czech Republic.
Introduction
The Silurian/Devonian boundary GSSP section at Klonk near Suchomasty has been revisited by drilling a shallow and fully cored borehole adjacent to the stratotype section. Both, the stratotype section and the core, were studied palynologically with respect to the exact position of the boundary in the framework of a multidisciplinary approach, including geochemistry and sedimentology.
Organic–walled microfossils from the stratotype section have been studied several times during the last 25 years. Chitinozoans were described by Paris et al. (1981) and distinct changes in the compostion of their assemblages served together with conodonts, graptolites and trilobites to define the system boundary. Acritarch and/or spore data have been only shortly discussed by Deunff (1980), Richardson (1984), Richardson et al. (1981), Le Hérissé et al. (1997) Le Hérissé (2002), and more recently also by Brocke et al. (2002). All these studies documented the lack of stratigraphically important acritarchs and spores at the GSSP.
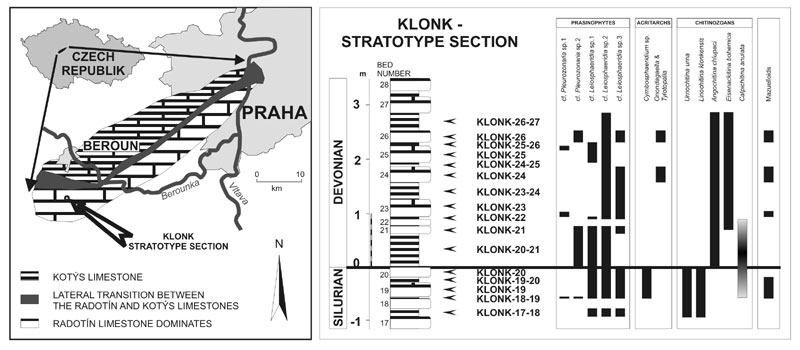
Recently, sixteen samples taken from the stratotype section (Figure 1. Klonk 17–18 to KLONK 26–27) have been studied for organic–walled microfossils. The samples were processed by standard palynological methods (HCl, HF treatment without oxidation). The received organic residue was separated into two fractions 63 µm or 40 µm and 10 µm respectively.
Palynology
Within the S/D boundary interval the distal marine palynological assemblages show a pronounced distribution pattern of prasinophytes, acritarchs, mazuelloids, scolecodonts and tubular filaments. The composition of the observed assemblages fits well with the generally accepted deeper outer shelf or even basinal depositional environment. Rare spores in the succeeding lower part of the Lochkovian Stage characterise a slowly increasing input of terrestrially derived organic matter in a shallowing environment during a suspected regressive phase. However, quantitative analysis of palynomorphs from the stratotype section (Figure 1) indicates a specific scenario around the Silurian/Devonian transition.
Figure 1. A. Facies distribution in the lower part of the Lower Devonian Lochkov Formation (modified after Chlupác, 1977, and Chlupác and Kukal, 1977). B. The Silurian/Devonian boundary interval in the GSSP at Klonk near Suchomasty with studied samples and the distribution of selected palynomorphs.
Prasinophytes of five basic morphotypes have been distinguished based on the character of the mesh–like ornamentation (cf. Pleurozonaria sp. 1 and cf. Pleurozonaria sp. 2) and/or thickness of the vesicle wall combined with the vesicle diameter (Leoisphaeridia sp. 1, Leoisphaeridia sp. 2 and Leoisphaeridia sp. 3). Both morphotypes of the genus Pleurozonaria occur together in one sample below the boundary (Figure 1, sample KLONK 18–19), but show differences in distribution in the lower Devonian (Figure 1).
Acritarchs are represented by six genera (Cymbosphaeridium spp., Diexallophasis sp., Micrhystridium spp., Multiplicisphaeridium sp. Onondagaella sp., Tylotopalla sp.). The genus Cymbosphaeridium Lister 1970 is common in the uppermost Silurian, however does not cross the Silurian/Devonian boundary. The genera Onondagaella Cramer 1966, emend. Playford 1977 and Tylotopalla Loeblich 1970 are very rare and in the stratotype section have been documented in Devonian only. Other genera are very sparsely distributed across the Silurian/Devonian boundary and have low stratigraphic potential.
Chitinozoans have been proved to be most indicative for stratigraphic purposes (Paris et al., 1981 and Paris, 1996). The Silurian/Devonian boundary can be fixed by the FAD of Angochitina chlupaci Paris and Laufeld, 1981 and the LAD of Linochitina klonkensis Paris and Laufeld, 1981 and Urnochitina urna (Eisenack, 1934). Further characteristic species for the Silurian/Devonian boundary interval are Eisenackitina bohemica (Eisenack, 1934) and Calpichitina annulata (Paris, 1981). C. annulata shows a distinct accumulation ("acme–zone") within the boundary interval (Figure 1).
Scolecodonts and tubular filaments (Porcatitubulus sp. and Laevitubulus sp.) have also been found sporadically. Phytodebris and the first spores occur later in the Lochkovian, reflecting more proximal environmental conditions. Altogether, these groups are scarce and could not be used for biostratigraphy.
Conclusions
The highest taxonomic diversity of palynomorphs has been documented in the uppermost Prídolí and in the lower part of the Lochkovian. The Silurian/Devonian boundary interval contains comparatively poorly diversified palynomorph assemblages, in which some groups are relatively rare (chitinozoans) or even totally missing (acritarchs, scolecodonts, certain prasinophytes, mazuelloids) in samples of the basal Lochkovian (Figure 1). The boundary interval is characterised by the dominance of thick– and/or thin–walled prasinophytes like as cf. Leiosphaeridia spp. and cf. Pleurozonaria spp. Such pattern of distribution could be connected with a transgressive pulse occurring just at the boundary, which was preceeded and also followed by regressions. Geochemical and sedimentological data fit well with these results. Comparable succession of palynological assemblages has been also observed in Lower–Middle Ordovician sequences in south–west China (Brocke, 1998).
Acknowledgements
The study was funded by the German Research Foundation (DFG) as part of the projects MA 1861/2–2 and BR 1943/3–1, and the Humboldt Foundation (V–8121/TSR/1007014). Thanks are due to the Research Centre Juelich (Dr. U. Mann) for drilling the borehole at Klonk and access to the core material.
References
Brocke, R. 1998. Palynomorpha (Acritarchen; Prasinophyceae, Chlorophyceae) aus dem Ordovizium der Yangtze–Platform, Sudwest–China. PhD Thesis, Technical University Berlin: 1–225.
Brocke, R., Wilde, V., Fatka, O. and Mann, M. 2002. Chitinozoa and acritarchs at the Silurian/Devonian boundary: Examples from the Barrandian area. 1st International Palaeontological Congress. Sydney.
Deunff, J. 1980. Le paléoplancton des Grés de Landévennec (Gedennien de la Rade de Brest, Finistère) étude biostratigraphique. Geobios, 13: 483–539.
Chlupác, I. 1977. Barrandian. In: Martinsson A. (ed.) The Silurian–Devonian boundary. IUGS Series A, 5: 84–95.
Chlupác, I. and Kukal, Z. 1977. The boundary stratotype at Klonk. In: Martinsson A. (ed.) The Silurian–Devonian boundary. IUGS Series A, 5: 96–109.
Le Hérissé, A. 2002. Paleoecology, biostratigraphy and biogeoegraphy of late Silurian to early Devonian acritarchs and prasinophycean phycomata in well A161, Western Libya, North Africa. Review of Palaeobotany and Palynology, 118(1–4): 359–395.
Le Hérissé, A., Gourvennec, R. and Wicander R. 1997. Biogeography of Late Silurian and Devonian acritarchs and prasinophytes. Review of Palaeobotany and Palynology, 98(1): 105–124.
Paris, F. 1996. Chitinozoan biostratigraphy and paleoecology. In: Jansonius, J. and McGregor, D.C. (eds.) Palynology: Principals and Applications, 2: 531–552.
Paris, F., Laufeld, S. and Chlupac, I. 1981. Chitinozoa of the Silurian–Devonian boundary stratotypes in Bohemia. Sveriges Geologiska Unders., Ser. Ca, 51: 1–29.
Richardson, J.B. 1984. Mid–Paleozoic palynology, facies and correlation. Procedings of 27th International Geological Congress, 1: 341–365.
Richardson, J.B., Rasul, M. and Al–Ameri, T. 1981. Acritarchs, miospores and correlation of the Ludlowian–Downtonian and Silurian–Devonian boundaries. Review of Palaeobotany and Palynology, 34: 209–224.
Received: February 15, 2003
Accepted: June 15, 2003