The distribution of Ostracoderms in the Mid-Ordovician of Laurentia and Gondwana
Ivan. J. Sansom1, M. Paul Smith1 and Philip J. C. Donoghue1
1 Lapworth Museum of Geology, School of Geography, Earth and Environmental Sciences, University of Birmingham, Birmingham, B15 2TT, U.K. E–mail: i.j.sansom@bham.ac.uk
Key words: Vertebrates. Gnathostomes. Agnathans. Conodonts. Palaeobiogeography.
Introduction
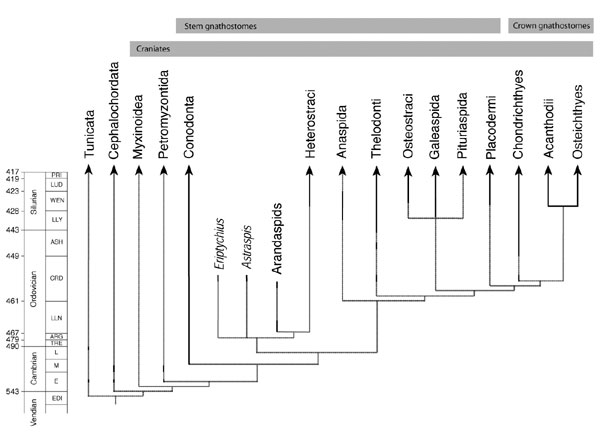
Recent discoveries have dramatically altered traditional views on the phylogeny, stratigraphic distribution and palaeobiogeography of Early Palaeozoic vertebrates (Donoghue et al., 2000; Sansom et al., 2001; Smith et al., 2002), and have led to a revised understanding of the origin, early evolution, and function of the early vertebrate skeleton (Donoghue and Sansom, 2002; Smith and Sansom, 2000). Not withstanding this, cladistic and ghost range analyses on the expanded dataset have identified large gaps in the stem-group gnathostome fossil record during the Ordovician (Smith et al., 2002; Figure 1). This interval is critical to our understanding of the origins and early evolution of the jawed vertebrates, one of the major innovations in the history of the vertebrate clade.
Palaeobiogeography
Two major patterns are apparent in the biogeographic data (Smith et al., 2002). Firstly, the majority of jawless fishes with dermoskeletal, plated ‘armour’ were highly endemic during Cambrian–Ordovician time, with arandaspids restricted to Gondwana, galeaspids to China, and anatolepids, astraspids and, possibly, heterostracans confined to Laurentia. The Laurentian groups began to disperse to other continental blocks as the ‘Old Red Sandstone continent’ amalgamated through a series of tectonic collisions. The second major pattern, in contrast, encompasses a number of microsquamous and naked, jawed and jawless primitive vertebrates such as conodonts, thelodonts, placoderms, chondrichthyans and acanthodians, which dispersed rapidly and crossed oceanic barriers to attain cosmopolitan distributions, although many have Laurentian origins. This is in marked contrast to previous studies that have suggested an "out-of-Gondwana" model (see the dispersal model of Elliott et al., 1991), although the comments of Young (in Webby et al., 2000) highlight the paucity of data available to Elliott et al.
Laurentian faunas
Cambro-Ordovician non-conodont vertebrates have been known from Laurentia since Walcott (1892) described two taxa from the Harding Sandstone of Colorado. Sporadic studies of shallow marine facies flanking the Transcontinental Arch identified the presence of a similar Caradoc fauna – dominated by the heterostracomorphs Astraspis and Eriptychius – in Wyoming (Darton, 1906), Oklahoma (Ossian and Halseth, 1976), subsurface Montana (Ross, 1957) and Ontario (Darby 1982). Work conducted by the authors and others has greatly expanded knowledge of Laurentian Early Palaeozoic faunas, both in terms of their phylogenetic diversity (Sansom et al., 1996, 2001), temporal occurrence (Smith et al., 1996, 2001; Clark et al., 1999) and spatial distribution (Sansom and Elliott 2002), together with an understanding of the palaeobiology of previously described forms (Sansom et al., 1997; Smith and Sansom, 1997).
Figure 1. The stratigraphic distribution and phylogenetic relationships of Lower Palaeozoic vertebrates. Solid lines indicate observed occurrences, dashed lines indicate inferred ghost ranges. Based upon Smith et al., 2002, Figure 4, with absolute ages taken from sources in Cooper (1999). The majority of lineages are predicted to have their first appearance in the Mid-Ordovician.
Ecologically, the faunas flanking the Transcontinental Arch are divided into a shoreface guild, characteristic of coarse siliciclastic sediments and the taxa Astraspis, Eriptychius and Skiichthys, whereas chondrichthyan and thelodont taxa (such as those described by Sansom et al., 1996), together with a host of unassigned forms, are found in quieter water, presumably inner shelf to lower shoreface sediments (see comments in Smith and Sansom, 1997). The investigation of the latter environment has provided the greatest yield of new taxa in the Laurentian Caradoc, and enabled the known record of major stem gnathostome clades such as the chondrichthyans and thelodonts from the Lower Silurian back into the Ordovician.
Gondwanan faunas
Gondwana has yielded pre-Silurian non-conodont vertebrates from four areas – Bolivia (Gagnier, 1993), the Precordillera and Eastern Cordillera of Argentina (Albanesi et al., 1995; Albanesi and Astini, 2002), and the Amadeus Basin of Australia (Ritchie and Gilbert Tomlinson, 1977; Young, 1997). Additionally, Young (in Webby et al., 2000) mentioned the possible presence of Ordovician ostracoderms in the Wiso and Canning Basins. The faunas described from Gondwana appear to be of lower diversity than those from Laurentia, being dominated by the arandaspids (Arandaspis and Sacabambaspis in Australia and Sacabambaspis alone in Bolivia and Argentina) in predominately shallow water shoreface environments.
Palaeoenvironmental settings
Initial literature and field studies suggest that both the Laurentian and Gondwanan mid-Ordovician faunas are represented in environments which are dominated by siliciclastic sedimentation in a shallow marine epicontinental setting. Diverse trace fossil assemblages associated with the occurrence of stem gnathostomes on both palaeocontinents suggest the repeated development of innershelf – shoreface – lagoonal parasequences. An invertebrate macrofauna dominated by molluscs (nautiloids and bivalves) is also characteristic of the Stairway Sandstone at Mount Watt, Northern Territories, Australia, and the main Harding Sandstone outcrop at Cañon City, Colorado, USA, both of which have a rich fauna of stem gnathostomes. The Larapinta Group of central Australia offers largely unrealised potential for the study of Ordovician vertebrates, as these conditions appear to persist throughout the Arenig–Caradoc range of the group, whereas the Harding Sandstone and allied units running parallel to the Laurentian Transcontinental Arch (Ross, 1976) is limited to the compressa to tenius conodont Chronozones of the middle Caradoc (Sansom and Smith, in press).
Conclusions
Although a burgeoning number of studies describing new vertebrate taxa from the mid-Ordovician have been published in the past decade, there is still considerable scope for further studies, particularly in Gondwana. These will provide a critical test of palaeogeographic and palaeoenvironmental models of early ostracoderm occurrences and dispersal.
References
Albanesi, G.L., Benedetto, J.L. and Gagnier, P.-Y. 1995. Sacabambaspis janvieri (Vertebrata) y conodontes del Llandeiliano temprano en la Formacion La Cantera, Precordillera de San Juan, Argentina. Boletin de la Academia Nacional de Ciencias, Cordoba, Argentina, 60: 519-543.
Albanesi G.L. and Astini, R.A. 2002. Faunas de conodontes y Sacabambaspis janvieri (Vertebrata) en el Ordovícico Medio de la Cordillera Oriental argentina: implicancias estratigráficas y paleobiogeográficas. In: L.A. Anzótegui, A.I. Lutz, and O.F. Gallego (Eds.) VIII Congreso Argentino de Paleontología y Bioestratigrafía, Corrientes, Resumen: 17a.
Clark, D.L., Sorenson, J.K., Ladd, A.N. and Freiheit, J.R. 1999. Probable microvertebrates, vertebrate-like fossils and weird things from the Wisconsin Ordovician. Journal of Paleontology, 73: 1201-1209.
Cooper, R.A. 1999. The Ordovician time scale – calibration of graptolite and conodont zones. Act Universitas Carolinae – Geologica, 43(1/2): 1-4.
Darby, D.G. 1982. The early vertebrate Astraspis, habitat based upon lithological association. Journal of Paleontology, 56: 1187-1196.
Darton, N.H. 1906. Fish remains in Ordovician rocks in Big Horn Mountains, Wyoming, with resume of Ordovician geology of the Northwest. Bulletin, Geological Society of America, 17: 541-566.
Donoghue, P.C.J. and Sansom, I.J. 2002. Origin and early evolution of vertebrate skeletonization. Microscopy, Research and Technique, 59: 352-372.
Donoghue, P.C.J., Forey, P.L. and Aldridge, R.J. 2000. Conodont affinity and chordate phylogeny. Biological Reviews, 75: 191-251.
Elliott, D.K., Blieck, A.R.M. and Gagnier, P.-Y. 1991. Ordovician vertebrates. In C.R. Barnes and S.H. Williams (Eds.) Advances in Ordovician Geology. Geological Survey of Canada, Paper 90-9: 93-106.
Gagnier, P.-Y. 1993. Sacabambaspis janvieri, vertébré Ordovicien de Bolivie: 1 analyse morphologique. Annales de Paléontologie, 79: 19-69.
Ossian, C.R. and Halseth, M.A. 1976. Discovery of Ordovician vertebrates in the Arbuckle Mountains of Oklahoma. Journal of Paleontology, 50: 773-777.
Ritchie, A. and Gilbert-Tomlinson, J. 1977. First Ordovician vertebrates from the southern Hemisphere. Alcheringa, 1: 351-368.
Ross, R.J. Jr. 1957. Ordovician fossils from wells in the Williston Basin, eastern Montana. United States Geological Survey, Bulletin 1021-M: 439-510.
Ross, R.J. Jr. 1976. Ordovician sedimentation in the western United States. In: Bassett, M.G. (Ed.), The Ordovician System: Proceedings of a Palaeontological Association Symposium, Birmingham, September 1974. University of Wales Press and National Museum of Wales, Cardiff: 73-105.
Sansom, I.J. and Elliott, D.K. 2002. Thelodonts from the Ordovician of Ontario. Journal of Vertebrate Paleontology, 22: 867-870.
Sansom, I.J., Smith, M.M. and Smith, M.P. 1996. Scales of thelodont and shark-like fishes from the Ordovician of Colorado. Nature, 379: 628-630.
Sansom, I.J., Smith, M.M. and Smith, M.P. 2001. The Ordovician radiation of vertebrates. In: P.E. Ahlberg (Ed.) Major events in early vertebrate evolution. Taylor and Francis, London: 156-171.
Sansom, I.J. and Smith, M.P. In press. Ordovician microvertebrates from the Bighorn Mountains of Wyoming. Palaeontology.
Sansom, I.J., Smith, M.P., Smith, M.M. and Turner, P. 1997. Astraspis – the anatomy and histology of an Ordovician fish. Palaeontology, 40: 625-643.
Smith, M.M. and Sansom, I.J. 1997. Exoskeletal micro-remains of an Ordovician fish from the Harding Sandstone of Colorado. Palaeontology, 40: 645-658.
Smith, M.M. and Sansom, I.J. 2000. Evolutionary origins of dentine in the fossil record of early vertebrates: diversity, development and function. In: M.F. Teaford, M.M. Smith and M.W.J. Ferguson (Eds.), Development, function and evolution of teeth. Cambridge University Press, Cambridge: 65–81.
Smith, M.P., Donoghue, P.C.J. and Sansom, I.J. 2002. Spatial and temporal diversification of Early Palaeozoic vertebrates. In: Crame, J.A. and Owen, A.W. (Eds.), Palaeobiogeography and biodiversity change: the Ordovician and Mesozoic-Cenozoic radiations. Special Publication of the Geological Society of London, 194: 69–83.
Smith, M.P., Sansom, I.J. and Cochrane, K.D. 2001. The Cambrian origin of vertebrates. In: P.E. Ahlberg (Ed.), Major events in early vertebrate evolution. Taylor and Francis, London: 67–84.
Smith, M.P., Sansom, I.J. and Repetski, J.E. 1996. Histology of the first fish. Nature, 380: 702-704.
Walcott, C.D. 1892: Preliminary notes on the discovery of a vertebrate fauna in Silurian (Ordovician) strata. Bulletin of the Geological Society of America, 3: 153-171.
Webby, B.D., Percival, I.G., Edgecombe, G.D., Cooper, R.A., Vandenberg, A.H.M., Pickett, J.W., Pojeta, J. Jr., Playford, G., Winchester-Seeto, T., Young, G.C., Zhen, Y.-Y., Nicoll, R.S., Ross, J.R.P. and Schallreuter, R. 2000. Ordovician palaeobiogeography of Australasia. Memoir of the Association of Australian Palaeontologists, 23: 63-126.
Young, G.C. 1997. Ordovician microvertebrate remains from the Amadeus Basin, central Australia. Journal of Vertebrate Paleontology, 17: 1-25.
Received: February 15, 2003
Accepted: June 15, 2003