Was the Yangtze Platform a refugium for graptolites during the Hirnantian (Late Ordovician) mass extinction?
Charles e. mitchell1, Michael j. Melchin2, h. David sheets3, Chen Xu4 and fan Jun–xuan4
1Department of Geology, SUNY at Buffalo, Buffalo, NY 14260. E–mail: cem@nsm.buffalo.edu
2 Department of Geology, St. Francis Xavier University, Antigonish, NS B2G 2W5.
3Department of Physics, Canisius College, Buffalo, NY 14208.
4Nanjing Institute of Geology and Paleontology, Academia Sinica, Nanjing, China.
Key words: Graptolites. Extinction. Hirnantian. Ordovician. Yangtze Platform.
Graptolites experienced near total extinction during the Hirnantian Mass Extinction (HME) in the latest Ordovician (Melchin and Mitchell, 1991; Koren’, 1991). This nadir followed by less than 3 My a mid–Ashgill peak diversity (measured as species/My) that was the greatest since the early Darriwilian, some 20 My earlier (Cooper, 1999). Using graphic correlation, we have compiled species ranges from the extraordinary Yangtze Platform faunas (Dicellograptus complexus to Parakidograptus acuminatus biozones, see Chen et al., 2000). The Yangtze fauna includes a high proportion of the graptolites species known to occur worldwide, as well as many endemic species. In addition, a range of graptolite biofacies are represented. Therefore, these data allow us to investigate the history of graptolite diversity changes within a finely resolved succession of intervals with equal duration (in My) that have been calibrated to graptolite chronozones and geochronometric ages (Cooper and Sadler in press; Melchin et al., in press).
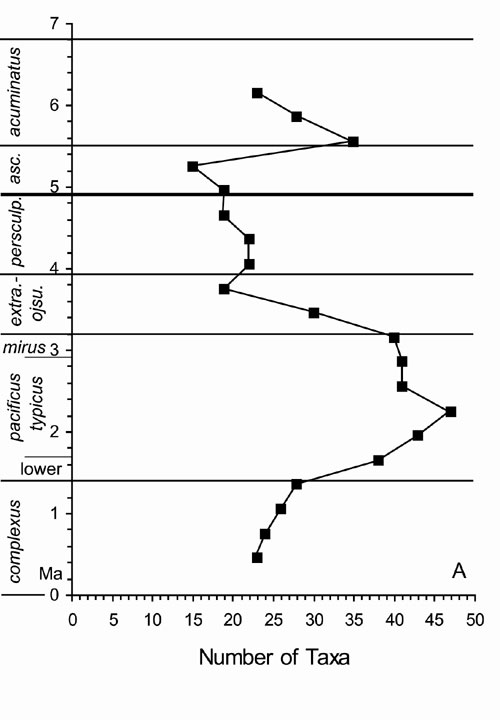
Graptolite species diversity (Figure 1) reached its zenith in the mid–P. pacificus Chron, following which it declined precipitously (irrespective of whether we use numbers of taxa or estimated mean standing diversity as our metric; see Foote, 2000). Compared to the deeper–water parts of the basin, this decline in diversity began slightly earlier (during the early P. pacificus Chron) in the shallower–water, more near–shore parts of the basin. Peak diversity also remained slightly lower and fell to much lower values in the near–shore part of the basin. Species diversity recovered slightly during the N. persculptus Chron but declined again in the late N. persculptus and early A. ascensus chrons.
Application of Capture–Mark–Recapture methods (see Connolly and Miller, 2001), including implementation of a unique bootstrapping method to obtain confidence limits on rates, facilitates testing for homogeneity of extinction and origination rates through the HME. The best fit model is the model that allows both extinction rate and origination rate to vary with time. Thus, we reject the hypothesis that graptolite evolutionary dynamics were constant through the late Ashgill to early Rhuddanian and that the HME might be a consequence of coincidental set of random extinctions. During all but the latest of the pre–Hirnantian part of the study period, estimated per–capita origination rates were low and extinction rates were lower still. Thus, the expansion of diversity in the Late Ordovician appears to be a result primarily of very low extinction risk. Bootstrapped confidence intervals on the rates of extinction per interval calculated within the CMR analysis (Figure 2) confirm that the formerly very low rates of extinction began t o rise in latest P. pacificus Chron as the formerly dominant Ordovician clades (climacograptids, orthograptids, and dicellograptids; the DDO fauna) experienced accelerated extinction. The HME peak was accompanied by a significant rise in origination rates (possibly with a very short lag) during the late N. extraordinarius–N. ojsuensis Chron Thus, rediversification of the graptolite fauna began within the interval of mass extinction, primarily by the appearance of numerous new normalograptid species. Origination rates rose significantly again in the A. ascensus Chron as the main graptolite diversity recovery began.
Figure 1. Graptolite species diversity, expressed in total number of taxa per 0.3 My interval, for the Yangtze region. Values are plotted at the mid-point of each interval for which they are calculated. Numerical time scale arbitrarily chosen as 0 My at base of D. complexus Chronozone (˜ 448 Ma).
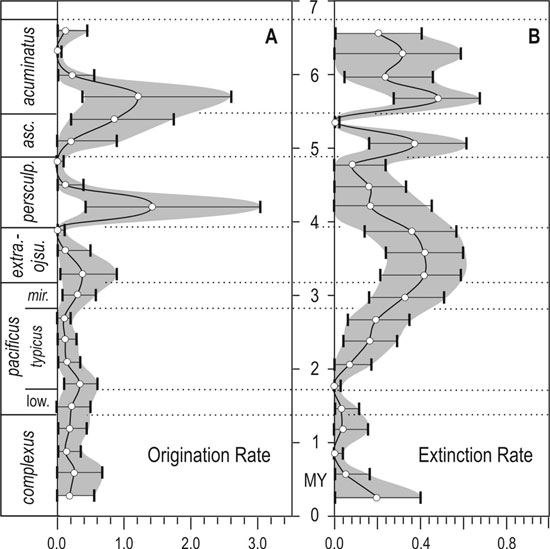
Figure 2. Origination and extinction rates with confidence intervals, calculated from Capture-Mark-Recapture analysis. Rates are per-taxon per interval, intervals are 0.3 My. Origination values are plotted at the beginning of each interval and extinction values at end of each interval for which they are calculated.
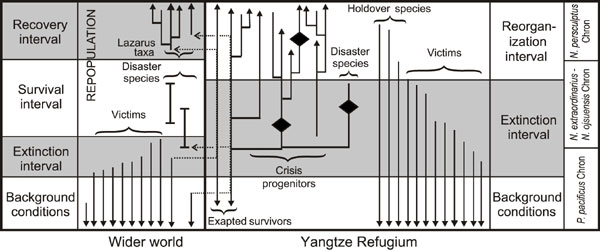
Figure 3. Conceptual model illustrating the patterns of graptolite species turn over in the Yangtze region and parts of the low latitude realm outside South China relative to the expected phases of turn over and ecological evolutionary character of species based on the model of Kauffman and Harries (1996).
Chen et al. (2003) have shown that there are strong geographic differences in the diversity patterns of graptolites during the Hirnantian Mass Extinction. High latitude regions experienced an earlier and more rapid decline in diversity than did low latitude regions. Yet, even after controlling for this effect, many graptolite species evidently survived considerably longer in the Yangtze Platform region than was the case elsewhere (Koren’ and Bjerreskov, 1999; Finney et al., 1999). At least five DDO holdover species (Paraorthograptus pacificus, P. brevispinus, Rectograptus abbreviatus, Anticostia uniformis, and Amplexograptus latus) persisted into the N. persculptus Chron in the Yangtze region, beyond the main interval of high extinction rates. A difference of a similar sort also exists in respect to several species of Normalograptus. For instance, N. normalis occurs in late Ashgill strata in many regions of the world, including the Yangtze Platform. In all these regions except Yangtze, it becomes extinct during the early Hirnantian, only to reappear as a Lazarus taxon with the return or graptolite–rich black shales in the late Hirnantian and early Rhuddanian. In the Yangtze Platform sections this species is present throughout the Hirnantian (Chen et al., in press). Finally, our current dynamical analyses suggest that the rise in origination rates among the mass extinction survivors in the Yangtze region followed immediately after the peak interval of mass extinction with essentially no interval ‘survival interval’ in which species diversity, E and O were all low (Figure 3). These patterns of graptolite turnover contrast markedly with theoretical expectations (e.g., Kauffman and Erwin, 1995; Kauffman and Harries, 1996) about mass extinction and recovery dynamics in which a substantial lag interval follows mass extinction and precedes diversity recovery. The delayed extinction of many graptolite species (including the unique presence of holdover taxa in the N. persculptus Chron), the persistence of species that elsewhere are Lazarus taxa, and the temporal overlap between the extinction of the DDO species and initial radiation of the normalograptids (i.e., the lack of a distinct survival phase separating the extinction and recovery phases), all suggest that the Yangtze Platform region experienced a different pattern of response to climate change than occurred in much of the rest of the low latitude realm during the Hirnantian Mass Extinction. In particular, we suggest these patterns indicate that the Yangtze Platform may have served as a refugium in which many species survived that elsewhere become extinct, and that it may have been from these lineages and this region that graptolite species diversity rebounded following the Hirnantian Mass Extinction. Further study of the taphonomy, biogeography, and chronostratigraphy of the Hirnantian as well as a detailed understanding of graptolite phyologeny will be required to test this hypothesis. We are continuing to pursue these goals.
References
Chen, X., Fan, J.x., Melchin, M.J. and Mitchell, C.E. In press. Graptolites of the Hirnantian Substage (latest Ordovician) from the Upper Yangtze Region, China. Palaeontology.
Chen, X., Melchin, M.J. Fan, J.x., and Mitchell, C.E. 2003. The Ashgillian graptolite fauna of the Yangtze regions and biogeographical distribution of diversity in the latest Ordovician. Bulletin de la Société Géologique de France, 175: 141-148.
Chen Xu, Rong Jia–yu, Mitchell, C. E., Harper, D. A. T., Fan Jun–xuan, Zhan Ren–bin, Zhang Yuan–dong, Li Rong–yu and Wang Yi. 2000. Late Ordovician to earliest Silurian graptolite and brachiopod biozonation from the Yangtze region, South China with a global correlation: Geological Magazine, 137: 623–650.
Connolly, S. R. and Miller, A. I. 2001. Joint estimation of sampling and turnover rates from fossil databases: capture–mark–recapture methods revisited. Paleobiology, 27: 751–767.
Cooper, R. A., 1999, Graptolites and the great Ordovician biodiversification event: In: P. Kraft, and Fatka, O., Eds., Quo vadis Ordovician? Acta Universitatis Carolinae – Geologica, 43: 441–442.
Cooper, R. A. and Sadler, P.M., In press. Chapter 15: Ordovician System. In: Gradstein, F.M. Ogg, J.G. and Smith, A.G., (Eds.). A Geologic Time Scale, Cambridge University Press.
Finney, S. C., Berry, W. B. N., Cooper, J. D., Ripperdan, R. L., Sweet, W. C., Jacobson, S. R., Soufiane, A., Achab, A., and Noble, P. J. 1999. Late Ordovician mass extinction: A new perspective from stratigraphic sections in central Nevada. Geology, 27: 215–218.
Foote, M., 2000, Origination and extinction components of taxonomic diversity: general problems. In: Erwin, D. H. and Wing, S. L., (Eds.). Deep Time: Paleobiology’s Perspective. Paleobiology, 26 (Supplement): 74–102.
Kauffman, E. G. and Erwin, D.H. 1995. Surviving mass extinctions. Geotimes, 14: 14–17.
Kauffman, E. G. and P. J. Harries. 1996. The importance of crisis progenitors in recovery from mass extinction. In: M. B. Hart (Ed.). Biotic Recovery from Mass Extinction Events. Geological Society Special Publication, 102: 25-39.
Koren’, T. N., 1991, Evolutionary crisis of the Ashgill graptolites: in Barnes, C. R. and Williams, S. H., Eds., Advances in Ordovician Geology, Geological Survey of Canada, paper 90–9, p. 157–164.
Koren, T. N., and M. Bjerreskov. 1999. The generative phase and the first radiation event in the Early Silurian monograptid history. Palaeogeography, Palaeoclimatology, Palaeoecology, 154: 3–9.
Melchin, M. J. and Mitchell C. E. 1991. Late Ordovician extinction in the Graptoloidea: in Barnes, C. R., and Williams, S. H., (Eds.). Advances in Ordovician Geology. Geological Survey of Canada, Paper 90–9, p. 143–156.
Melchin, M. J., Cooper, R. A. and Sadler, P. M. In press. Chapter 16: Silurian System. In: Gradstein, F.M. Ogg, J.G. and Smith A.G. (Eds.). A Geologic Time Scale, Cambridge University Press
Received: February 15, 2003
Accepted: June 15, 2003