Early Ordovician paleogeography of the Korean Peninsula
Sang Min Lee1 and Duck K. Choi1
1 School of Earth and Environmental Sciences, NS80, Seoul National University, Seoul 151–742, Korea. E–mail: dkchoi@snu.ac.kr
Key words: Trilobites. Paleogeography. Ordovician. North China. Korea.
Introduction
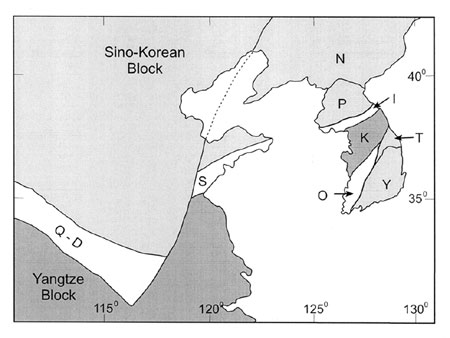
The Cambrian-Ordovician sedimentary rocks in southern Korea, the Choson Supergroup, are exposed in the Taebaeksan Basin, which occupies the mid-eastern part of the Korean peninsula (Figure 1). The Choson Supergroup consists predominantly of carbonate with lesser amounts of sandstone and shale, and is subdivided into the Taebaek, Yongwol, Yongtan, Pyongchang, and Mungyong groups (Choi, 1998a). Of these, the Taebaek and Yongwol groups are regionally extensive and fossiliferous, and hence are considered to represent the early Paleozoic environments and faunas of Korea. Kobayashi (1967) noted that the Cambrian faunas of the Taebaek Group belong to the Hwangho (North China) faunal province, which is characterized by endemic taxa inhabiting a shallow marine environment, whereas those of the Yongwol Group are comparable to the Jiangnan (South China) fauna, which is dominated by cosmopolitan and pelagic forms indicating a deeper–water oceanic setting. On the other hand, Kobayashi (1969) showed that the Ordovician cephalopod faunas of Korea and North China are distinct from those of South China. These faunal characteristics between North and South China led to suggest that the Sino–Korean and Yangtze blocks were separated during much of the Paleozoic (Burrett, 1973). However, detailed continental configuration around the Korean peninsula was always obscured in the global–scale paleogeographic maps (e.g., Scotese and McKerrow, 1991), due to the lack of reliable paleogeographic information within the Korean peninsula. The Korean peninsula was either included in the Sino–Korean block (Laurie and Burrett, 1992) or simply divided into the two parts, placing North and South Korea to the Sino–Korean and Yangtze blocks respectively (Watson et al., 1987). In this paper we present an early Paleozoic paleogeographic reconstruction around the Korean peninsula, based chiefly on the recently accumulated paleontological data from the Taebaeksan basin.
Ordovician trilobite faunal successions in Korea
Two independent biostratigraphic schemes have been employed for the Cambrian–Ordovician sequences of the Taebaeksan basin (Kobayashi, 1966). In the Ordovician, five trilobite biozones or faunas were recognized in the Taebaek Group, whereas four zones were established in the Yongwol Group. The Taebaek Group comprises the Pseudokainella, Asaphellus, Protopliomerops, Kayseraspis, and Dolerobasilicus zones, while the Yongwol Group includes the Yosimuraspis, Kainella, Shumardia, and Kayseraspis zones (Choi et al., 2001).
Figure 1. Tectonic division of the Korean peninsula and adjacent areas. (I, Imjingang belt; K, Kyonggi massif; N, Nangrim massif; O, Okchon belt; P, Pyongnam basin; Q-D, Qinling-Dabie belt; S, Sulu belt; T, Taebaeksan basin; Y, Yongnam massif).
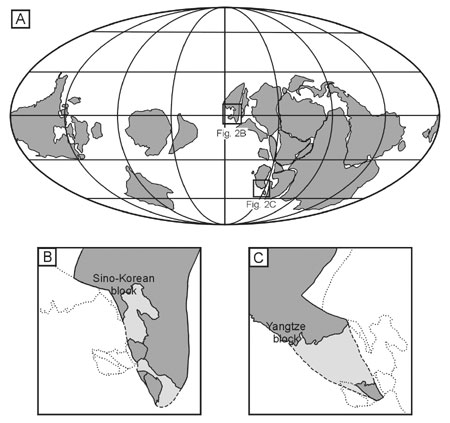
Figure 2. Early Ordovician paleogeographic maps. A. Tremadocian paleogeographic reconstruction, modified from Scotese and McKerrow (1991, figure 2); B. Sino-Korean block; C. Yangtze block. Note that the Yongnam Massif has been rotated clockwise considerably since the collision between the Sino-Korean and the Yangtze block. The present coastline and the tectonic divisions shown in Figure 1 are indicated by dotted lines in B and C.
The Pseudokainella Zone was established based on the sole occurrence of Pseudokainella iwayai (Kobayashi, 1966). The upper Tremadocian Asaphellus Zone yields Asaphellus, Kainella, and Hystricurus. The succeeding Protopliomerops Zone also contains a trilobite fauna of late Tremadocian age, represented by Shumardia, Apatokephalus, Dikelokephalina, Protopliomerops, and Seisonia. The Kayseraspis Zone comprises Hystricurus, Kayseraspis, Illaenus, Dikelokephalina, Chosenia, Asaphopsoides and Koraipsis. The Darriwilian Dolerobasilicus fauna includes Dolerobasilicus yokusensis, Basiliella kawasakii, B. typicalis, and Ptychopyge dongjeomensis (Lee and Choi, 1992).
The Yosimuraspis Zone yields the earliest Tremadocian trilobites including Yosimuraspis, Jujuyaspis, Elkanaspis, and pliomerid gen. and sp. indeterminate (Kim and Choi, 2000a). The middle Tremadocian Kainella Zone is composed of Kainella, Leiostegium, and agnostid gen. and sp. indeterminate (Kim and Choi, 2000b). The succeeding Shumardia Zone contains a diverse trilobite assemblage of late Tremadocian age, composed of Micragnostus, Shumardia, Apatokephalus, Hystricurus, Dikelokephalina, Asaphellus, Hukasawaia, Koraipsis, and pliomerid gen. and sp. indeterminate (Choi et al., 1994). The Kayseraspis fauna consists of Kayseraspis and Asaphopsoides, bellerophontoid gastropods, and ostracods (Choi, 1998b).
Biogeographic consideration of the Ordovician trilobite faunas
In the Taebaeksan basin, trilobites are among the most dominant fossil groups in the Early Ordovician, whereas they become less important in the Middle Ordovician faunal assemblages. Accordingly, discussion will be mainly focused on the Early Ordovician trilobite faunas. The trilobite faunas are characterized by frequent occurrence of pandemic polymeroid trilobites including Jujuyaspis, Leiostegium, Asaphellus, Shumardia, Apatokephalus, Hystricurus, Protopliomerops, Illaenus, Asaphopsoides, and Kayseraspis. The trilobite taxa with restricted distribution are Yosimuraspis, Dikelokephalina, Koraipsis, Hukasawaia, Seisonia, and Chosenia. These Early Ordovician trilobite faunas of the Taebaeksan basin show a close affinity to those of North China: most of these genera listed above were documented in North China. In addition, closely comparable Ordovician lithostratigraphic succession in North China implies that the Taebaeksan basin was connected through contiguous shallow waters to North China, occupying a same biogeographic province along the margins of the Sino–Korean block (Figure 2). The Sino–Korean faunas display close biogeographic ties with the Australian ones in sharing a number of trilobite genera, often at the species level (Shergold, 1991), whereas the South Chinese faunas lack some of the biogeographically significant taxa such as Jujuyaspis, Kainella, Koraipsis, and Kayseraspis among others. Interestingly, the biogeographic connection of these Early Ordovician Sino–Korean faunas may be extended to South America and North America through the occurrence of Jujuyaspis, Kainella, Leiostegium, and Kayseraspis. The co–occurrence of a fair number of trilobite genera in the peri–Gondwanaland and Laurentia in the Early Ordovician may be attributable to the disposition of these continents in the equatorial region (Scotese and McKerrow, 1991), thereby facilitating animals to cross the oceans via equatorial currents.
Paleogeographic reconstruction
In the early Paleozoic, the Korean peninsula must have been divided into three major parts: i.e., Nangrim, Yongnam, and Kyonggi massifs. The Nangrim and Yongnam massifs were part of the Sino–Korean block, whereas the Kyonggi massif belonged to the Yangtze block (Figures 1 and 2; Cluzel et al., 1991). The Cambro–Ordovician shallow marine sediments were accumulated on the margin (Pyongnam and Taebaeksan basins) of the Nangrim and Yongnam massifs, which were further connected to cratonic region of North China. The Taebaeksan basin was considered as a miogeoclinal setting fringing the Yongnam massif (Choi et al., 2001), which was part of the Sino–Korean block during the Paleozoic. These massifs were amalgamated to form much of the present Korean peninsula during the Late Permian to Early Triassic when the Sino–Korean and Yangtze blocks collided (Yin and Nie, 1993; Meng and Zhang, 1999).
References
Burrett, C.F. 1973. Ordovician biogeography and continental drift. Palaeogeography, Palaeoclimatology, Palaeoecology, 13: 161–201.
Choi, D.K. 1998a. The Yongwol Group (Cambrian–Ordovician) redefined: a proposal for the stratigraphic nomenclature of the Choson Supergroup. Geosciences Journal, 2: 220–234.
Choi, D.K. 1998b. An Early Ordovician trilobite faunule from the Choson Supergroup, Maepo, Tanyang area, Korea. Geosciences Journal, 2: 220–234.
Choi, D.K., Park, G.H. and Kim, D.H. 1994. Tremadocian trilobites from the Mungog Formation, Yeongweol area, Korea. Journal of the Paleontological Society of Korea, 10: 209–226.
Choi, D.K., Kim, D.H. and Sohn, J.W. 2001. Ordovician trilobite faunas and depositional history of the Taebaeksan Basin, Korea: implications for palaeogeography. Alcheringa, 25: 53–68.
Cluzel, D., Lee, B.J. and Cadet, J.P. 1991. Indosinian dextral ductile fault system and synkinematic plutonism in the southwest of the Ogcheon belt (South Korea). Tectonophysics, 194: 131–151.
Kim, D.H. and Choi, D.K. 2000a. Jujuyaspis and associated trilobites from the Mungok Formation (Early Ordovician), Yongwol, Korea. Journal of Paleontology, 74: 1031–1042.
Kim, D.H. and Choi, D.K. 2000b. Lithostratigraphy and biostratigraphy of the Mungok Formation (Lower Ordovician), Yongwol, Korea. Geosciences Journal, 4: 301–311.
Kobayashi, T. 1966. The Cambro–Ordovician formations and faunas of South Korea, Part X, Stratigraphy of the Chosen Group in Korea and South Manchuria and its relation to the Cambro–Ordovician formations of other areas, Section A, The Chosen Group of South Korea. Journal of the Faculty of Science, University of Tokyo, Section II, 16: 1–84.
Kobayashi, T. 1967. The Cambro–Ordovician formations and faunas of South Korea, Part X, Stratigraphy of the Chosen Group in Korea and South Manchuria and its relation to the Cambro–Ordovician formations of other areas, Section C, The Cambrian of eastern Asia and other parts of the continent. Journal of the Faculty of Science, University of Tokyo, Section II, 16: 381–534.
Kobayashi, T. 1969. The Cambro–Ordovician formations and faunas of South Korea, Part X, Stratigraphy of the Chosen Group in Korea and South Manchuria and its relation to the Cambro–Ordovician formations of other areas, Section D, The Ordovician of eastern Asia and other parts of the continent. Journal of the Faculty of Science, University of Tokyo, Section II, 17: 163–316.
Laurie, J.R. and Burrett, C. 1992. Biogeographic significance of Ordovician brachiopods from Thailand and Malaysia. Journal of Paleontology, 66: 16–23.
Lee, D.C. and Choi, D.K. 1992. Reappraisal of the Middle Ordovician trilobites from the Jigunsan Formation, Korea. Journal of the Geological Society of Korea, 28: 167–183.
Meng, Q.R. and Zhang, G.W. 1999. Timing of collision of the North and South China blocks: controversy and reconciliation. Geology, 27: 123–126.
Scotese, C.R. and McKerrow, W.S. 1991. Ordovician plate tectonic reconstructions. In: Barnes, C.R. and Williams, S.H. (Eds.), Advances in Ordovician Geology, Geological Survey of Canada, Paper 90–9: 271–282.
Shergold, J.H. 1991. The Pacoota Sandstone, Amadeus Basin, Northern Territory: stratigraphy and palaeontology. Bulletin of the Bureau of Mineral Resources of Australia, 237: 1–93.
Watson, M.P., Hayward, A.B., Parkinson, D.N. and Zhang, Z.M. 1987. Plate tectonic history, basin development and petroleum source rock deposition onshore China. Marine and Petroleum Geology, 4: 205–225.
Yin, A. and Nie, S. 1993. An indentation model for the North and South China collision and development of the Tan–Lu and Honam fault systems, eastern Asia. Tectonics, 12: 801–813.
Received: February 15, 2003
Accepted: June 15, 2003