Notes on the Ordovician trilobite radiation in the South China Block
Zhou Zhiyi1
1 Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Chi–Ming–Ssu, Nanjing 210008, China. E–mail: zyizhou@jlonline.com
Key words: Trilobites. Faunas. Evolution. Ordovician. China
Introduction
Adrain et al. (1998, in press) grouped Ordovician trilobites into two major faunas, the Ibex Fauna and Whiterock Fauna. The former had high diversity in the Early Ordovician but then declined rapidly and did not survive the end Ordovician mass extinction. The latter diversified rapidly during the Middle Ordovician radiation and gave rise to the Silurian Fauna then. Therefore, as they pointed out, the Whiterock Fauna had played an important role in the rise to dominance of the Sepkoski’s (1981) Palaeozoic Evolutionary Fauna. However, the time of onset and the environmental pattern of the trilobite radiation may vary palaeogeographically. For examples, the Whiterock Fauna became dominant and occupied environmental niches of shelf to upper slope in the early Arenig of Avalonia (Owen and McCormick, in press), but in Laurentia it began to radiate first in the platform–margin environments during the late Arenig or early Whiterockian (Adrain et al., 1998, in press).
The South China Block was situated in low latitude zones along the western margin of Gondwana during the Ordovician (Cocks, 2001). Ordovician rocks are extensively exposed there with records of complete sequences of various fossil groups. Trilobites are much diverse and faunal differentiation along bathymetric gradients and in response to biofacies has recently been explored (e.g., Zhou et al., 1999, 2001; Yuan et al., 2000), providing much significant information for investigating the radiation of the Whiterock Fauna in relation to environments.
Faunal replacement through the Ordovician –a review
As recorded by the related literature (e.g., Lu, 1975), faunal composition changed rapidly stage by stage through the Ordovician in South China. Trilobite clades that dominate each of successive chronostratigraphic units are Agnostina, Dikelokephalinidae, Ceratopygidae and Olenidae (Tremadocian); Asaphidae and Nileidae (late Early Ordovician); Asaphidae, Cyclopygidae and Trinucleidae (early Middle Ordovician); Asaphidae, Illaenidae, Cyclopygidae and Raphiophoridae (Darriwilian); Cyclopygidae, Aulacopleuroidea, Illaenidae, Raphiophoridae, Cheiruridae, Asaphidae and Agnostina (Caradoc–middle Ashgill); Dalmanitidae (Hirnantian). The list indicates that dominant components of the Early Ordovician all belong to the Ibex Fauna, but from the Middle Ordovician on most of them, except Asaphidae and Agnostida, are those of the Whiterock Fauna.
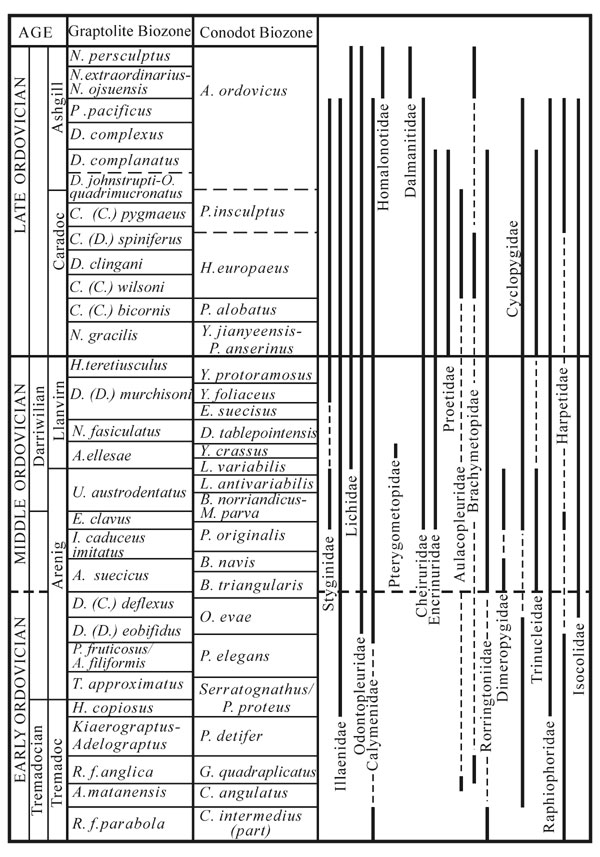
Figure 1. Chronostratigraphic distribution of the Whiterock Fauna families in the Ordovician of the South China Block. Correlation between graptolite and conodont biozones is after Chen et al. (2001), Wang et al. (1996) and Wang and Wang (2001) with modifications.
Composition and temporal distribution of the Whiterock Fauna
Altogether 21 from 24 trilobite clades that assigned to the Whiterock Fauna by Adrain et al. (in press) occur in the Ordovician of South China (Figure 1) (note that, following Adrain (personal communication, November, 2002), taxa of Phillipsinellidae, Celmidae and Staurocephalidae are here referred respectively to Styginidae, Dimeropygidae and Encrinuridae). There was a steady climb in the family diversity through most of the Ordovician followed by a sudden decrease in the Hirnantian. However, it is only the Hirnantian trilobite fauna that consists totally of elements of the Whiterock Fauna (Figure 2). Trilobites that survived the first–phase of the end–Ordovician mass extinction in South China are largely immigrants from high–latitude Gondwana, comprising genera such as Dalmanitina (Dalmanitidae), Eoleonaspis (Odontopleuridae) and Platycoryphe (Homalonotidae).
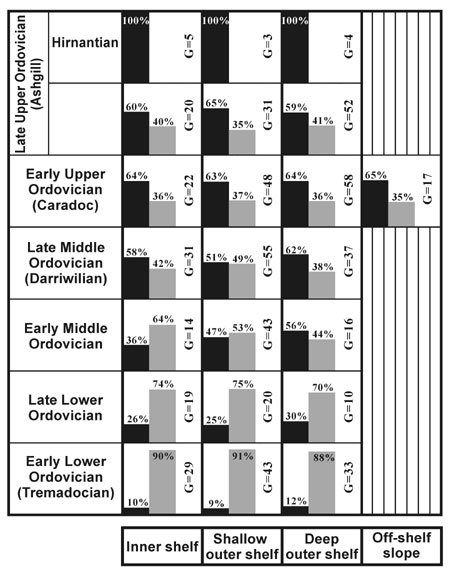
Figure 2. Histograms showing percentages of Ordovician trilobite genera belonging respectively to the Ibex Fauna (grey) and Whiterock Fauna (black) for each of 7 sampled intervals along an environmental gradient of the South China Block.
Nevertheless, numerical statistics of trilobite genera (Zhou, in press) indicate that it was until the late early Middle Ordovician (interval of the Exigraptus clavus graptolite Biozone), or the late Fennian in terms of the British chronostratigraphic standard (see Fortey et al., 2000), when taxa of the Whiterock Fauna first rivaled those of the Ibex Fauna. It means that the rise to dominance of the Whiterock Fauna, or the onset of the Ordovician trilobite radiation, was later in the South China Block than in Avalonia (see Owen and McCormick, in press) and Laurentia and Baltica (see Adrain et al., 1998, in press).
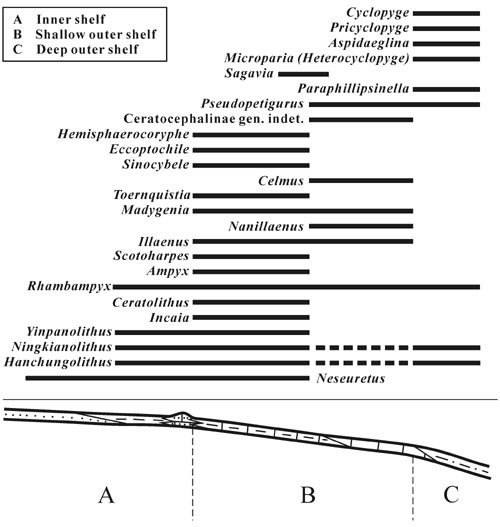
Figure 3. Model showing palaeogeographic distribution of most of the early Middle Ordovician Whiterock Fauna trilobites in the South China Block.
Among families listed in Figure 1, Cyclopygidae, Trinucleidae, Raphiophoridae, Illaenidae, Calymenidae and Cheiruridae are characteristic of the Whiterock Fauna in South China. They are not only much more diverse but also have more frequent occurrences. Many of the genera are typical of peri–Gondwana (Zhou and Dean, 1989).
Ordovician trilobite radiation in relation to environments
Ordovician trilobite faunas display a progressive on–shelf to off–shelf transition in composition and diversity from present–day west to either east or north of the South China Block. On the basis of data from the previous literature and recent collections made from 36 measured Ordovician sections along a bathymetric gradient in South China by Zhou Zhiyi, Zhou Zhiqiang, Yuan Wenwei and Samuel Turvey, more than 30 trilobite biofacies or associations are recognized. Some of them mark inner–shelf environments, while most have either a shallow or a deep outer–shelf setting. Only two off–shelf slope trilobite biofacies (Cyclopygid and Cyclopygid–Girvanopyge) were developed in the southern marginal area of the block during the middle Caradoc (Zhou et al., 1999), a period when the Ordovician climax transgression took place. Data for representative trilobite faunas that are associated with different biofacies are synthesized in Figure 2, which indicates that elements of the Whiterock Fauna were distributed and proportionally increased steadily through the Ordovician over the whole environmental spectrum. However, during the early Middle Ordovician members of the Whiterock Fauna first exhibited a higher representation in outer–shelf faunas and, especially, they first became predominant over those of the Ibex Fauna in deep outer–shelf environments. The Whiterock Fauna subsequently developed further and then dominated all environments from the Darriwilian onwards.
The early Middle Ordovician palaeogeography of the South China Block has been outlined by Zhang et al. (2002). Inner–shelf deposits (arenite, argillite, biocalcarenite and biocalcirudite) were diachronously distributed in the west and shallow outer–shelf rocks (mainly biocalcilutite and argillite) in the central and north of the block, with a complete trilobite record throughout the interval from the A. suecicus to E. clavus graptolite biozones. The deep inner–shelf fauna is well represented by the trilobites described from Chengkou, Chongqing (Lu and Chang, 1974). The shallow outer–shelf trilobites have been recorded in western Hubei and Hunan, southern Shaanxi (Turvey and Zhou, 2002; Turvey, 2002) and southern Anhui (Yuan, 2002). Trilobite–bearing, deep outer–shelf beds (argillite and arenite) are sporadically exposed, and collections were only made from the interval of E. clavus–U. austrodentatus biozones at Changde, northwestern Hunan (Turvey, 2002) and Mianxian, southern Shaanxi (personal collection). Accordingly, the environmental setting of genera of the Whiterock Fauna that occurred during the onset of the trilobite radiation is shown in Figure 3. Following Adrain et al. (in press), some of them belong to clades that were widespread in the early Middle Ordovician (Illaenidae, Harpetidae, Raphiophoridae), and some were linked with mid– to low–latitude radiating groups, such as ceratocephaline Odontopleuridae, Dimeropygidae, Encrinuridae, Rorringtonidae and Styginidae. However, of most of these families each contains only a single genus. As a whole, at the beginning of the radiation in South China, the Whiterock Fauna was characterized by the proliferation of typical Gondwana forms, including reedocalymenine calymenids, eccoptochiline cheirurids, trinucleids and cyclopygids. In general, trinuceids prevailed in the deep inner and shallow outer shelf and cyclopygids held a dominant position in the deep outer shelf environments. Both of the trilobite clades were only restricted to the Ordovician and had their early diversification centered in high latitude zones of Gondwana (Adain et al., in press; Fortey, in press).
Cyclopygidae is a unique trilobite group, with a mesopelagic mode of life (Fortey, 1985). It is diagnostic of an Ordovician adaptive radiation (Fortey, in press). Genera of this family inhabited different depth levels in the water column (Zhou et al., 1994), in ecological niches that had never been occupied by the Cambrian trilobite faunas. Again, the diversification of Cyclopygidae and the establishment of the Cyclopygid Biofacies in South China were initiated during the late early Middle Ordovician.
Conclusions
Members of the Whiterock Fauna spread across whole of the environmental spectrum and in general increased steadily through the Ordovician of the South China Block. The Whiterock Fauna first exhibited a higher representation in shallow outer–shelf faunas in the early Middle Ordovician, but rose to dominance over those of the Ibex Fauna first in deep outer–shelf environments during the late early Middle Ordovician. Therefore, the Ordovician trilobite radiation commenced in the block not only much later than in high–latitude Gondwana but also slightly later than in low–latitude Laurentia. In South China genera of the Whiterock Fauna were largely linked with the peri–Gondwana faunas, and the onset of the Ordovician trilobite radiation was marked by the proliferation of Trinucleidae and Cyclopygidae, both of which had their early diversification centered in high latitude zones of Gondwana. From the Darriwilian onwards elements of the Whiterock Fauna had dominated all environments, but it was not until the Hirnantian that they comprised the whole fauna.
Acknowledgements
The research was supported by the Special Funds for Major State Basic Research Program (Nº G2000077700) and the National Natural Science Foundation of China (No. 40272001). Richard Fortey, Jonatham Adrain and Beatriz Waisfeld are acknowledged for constructive suggestions. Field assistance was provided by Zhou Zhiqiang, Yuan Wenwei and Samuel Turvey.
References
Adrain, J. M., Fortey, R. A. and Westrop, S. R. 1998. Post–Cambrian trilobite diversity and evolutionary faunas. Science, 280 (5371): 1922–1925.
Adrain, J. M., Westrop, S. R. and Fortey, R. A. In press. Trilobites: Global Patterns. In: Webby, B. D., Droser, M. L. Paris, F. and Percival, I. G. (Eds.), The Great Ordovician Biodiversification Event. Columbia University Press.
Cocks, L.R.M. 2001. Ordovician and Silurian global geography. Journal of the Geological Society, London 158(2): 197–210.
Chen Xu, Zhou Zhiyi, Rong Jiayu and Li Jun 2001. Ordovician series and stages in Chinese stratigraphy: steps toward a global usage. Alcheringa, 25(2): 131–141.
Fortey, R.A. 1985. Pelagic trilobites as an example of deducing the life habits of extinct arthropods. Transactions of the Royal Society of Edinburgh: Earth Sciences, 76(2–3): 219–230.
Fortey, R.A., Harper, D.A.T., Ingham, J.K., Owen,A.W., Parkes, M.A., Rushton, A.W.A. and Woodcock, N.H. 2000. A revised correlation of Ordovician rocks in the British Isles. The Geological Society Special Report 24, The Geological Society of London: 1–83.
Fortey, R. A. In press. Adaptive deployment in Ordovician trilobites. In: Webby, B. D., Droser, M. L. Paris, F. and Percival, I. G. (Eds.), The Great Ordovician Biodiversification Event. Columbia University Press.
Lu Yanhao, 1975. Ordovician trilobite faunas of central and southwestern China. Palaeontologica Sinica, New Series B 11: 1–463 (in Chinese and English).
Lu Yanhao and Chang Wentang 1974. Ordovician trilobites. In: Nanjing Institute of Geology and Palaeontology, Academia Sinica (ed.), A handbook of stratigraphy and palaeontology in Southwest China. Science Press, Beijing: 124–136 (in Chinese).
Owen, A. W. and McCormick, T. In press. Trilobites: Regional patterns in the Anglo–Welsh sector of Avolonia. In: Webby, B. D., Droser, M. L. Paris, F. and Percival, I. G. (Eds.), The Great Ordovician Biodiversification Event. Columbia University Press.
Sepkoski, J. J., Jr. 1981. A factor analytic description of the Phanerozoic marine fossil record. Paleobiology, 7(1): 36–53.
Turvey, S.T. and Zhou Zhiyi 2002. Arenig trilobite associations of Daping, Yichang, Hubei, South China. Acta Palaeontologica Sinica, 41(1): 10–18.
Turvey, S.T. 2002. Early Ordovician (Arenig) trilobites of the South China Plate: taxonomy, palaeoecology and palaeobiogeography. PHD thesis of Department of Earth Sciences & St. John’s College, University of Oxford: 1–422.
Wang Xiaofeng and Wang Chuanshang 2001. Tremadocian (Ordovician) graptolite diversification events in China. Alcheringa, 25(2): 155–168.
Wang Zhihao, Bergström, S.G. and Lane, H.R. 1996. Conodont provinces and biostratigraphy in Ordovician of China. Acta Palaeontologica Sinica 35(1): 26–59.
Yuan Wenwei, Zhou Zhiyi, Zhang Junming, Zhou Zhiqiang, Sun Xiaowen and Zhou Tianmei 2000. Tremadocian trilobite biofacies in western Hunan–Hubei. Journal of Stratigraphy, 24 (4): 275–282 (in Chinese with English abstract).
Yuan Wenwei 2002. Ontogeny, Heterochrony and Phylogeny of some Ordovician and Silurian trilobites from Yangtze Region. PHD thesis of Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences: 1–124 (in Chinese with English summary).
Zhang Yunbai, Zhou Zhiyi and Zhang Junming 2002. Sedimentary differentiation during the latest Early Ordovician–earliest Darriwilian in the Yangtze Block. Journal of Stratigraphy, 26 (4): 302–314 (in Chinese with English abstract).
Zhou Zhiyi and Dean, W. T. 1989. Trilobite evidence for Gondwanaland in east Asia during the Ordovician. Journal of Southeast Asian Earth Sciences, 3 (1–4): 131–140.
Zhou Zhiyi, McNamara, K.J., Yuan Wenwei and Zhang Tairong 1994. Cyclopygid trilobites from the Ordovician of northeastern Tarim, Northwest China. Records of the Western Australian Museum, 16 (4): 593–622.
Zhou Zhiyi, Zhou Zhiqiang and Yuan Wenwei 1999. Middle Caradoc trilobite biofacies of western Hubei and Hunan, South China. Acta Universitatis Carolinae, Geologica, 43 (1/2): 385–388.
Zhou Zhiyi, Zhou Zhiqiang and Yuan Wenwei 2001. Llanvirn–early Caradoc biofacies of western Hubei and Hunan, China. Alcheringa, 25(1): 69–86.
Zhou Zhiyi. In press. Trilobites: Regional patterns in South China. In: Webby, B. D., Droser, M. L. Paris, F. and Percival, I. G. (Eds.), The Great Ordovician Biodiversification Event. Columbia University Press.
Received: February 15, 2003
Accepted: June 15, 2003