Preliminary investigation on Early to Middle Ordovician brachiopod biodiversity of South China
Zhan Ren-bin1 and Rong Jia-yu1
1 Nanjing Institute of Geology and Palaeontology, CAS, Nanjing 210008, The People’s Republic of China. State Key Laboratory of Palaeobiology and Stratigraphy (NIGPAS). E–mail: rbzhan@nigpas.ac.cn
Key words: Brachiopods. Biodiversity. Evolution. Ordovician. China.
Introduction
The Ordovician Period encompasses one of the most profound evolutionary radiations (Sepkoski, 1995), which lasted for several millions of years (Droser and Sheehan, 1995) and was manifested as three diversity maxima from Middle to Late Ordovician (Webby, 2000). Brachiopoda, an important component of the Paleozoic Evolutionary Fauna, was very abundant in Early Paleozoic of South China. Since 1970s, a number of systematic works on Ordovician brachiopods of South China have been published. Most recently, all Ordovician and Silurian brachiopods of China are reviewed, so we can add Chinese data into international discussions including "the great Ordovician biodiversity event".
From our preliminary revision, the Ordovician radiation is well-marked in the fossil record of South China. The first diversity maximum of Ordovician brachiopods of South China is in late Early Ordovician, i.e., late early Arenig, while the first diversity increase of international trend commenced about the beginning of the Middle Ordovician, and reached a peak in the early Darriwilian (Webby, 2000). The same trend is also true to bivalves that experienced the most fundamental radiation in Early Ordovician (Cope and Babin, 1999). From Tremadoc to Llanvirn, the diversity changes of brachiopods in South China were manifested not only by the number of systematic taxa but also by the complicating paleoecology.
Early to Middle Ordovician rocks and brachiopods of South China
During Early to Middle Ordovician, three major stratigraphical regions (Yangtze, Jiangnan and Huanan Stratigraphic Regions) developed on South China Paleoplate corresponding to different paleogeographic conditions (Yangtze Platform, Jiangnan Slope and Zhujiang Basin) (Rong and Chen, 1987). The Yangtze Platform was subdivided into Upper and Lower parts, on which carbonate rocks (e.g., Nantsinkuan and Hunghuayuan formations) were developed from Tremadoc to early Arenig, and clastic deposits were accumulated during most of the Middle Ordovician time (e.g., Meitan Formation). The Early-Middle Ordovician sequence, generally several hundreds of meters thick, is usually continuous but the lithofacies are quite variable on the Yangtze Platform (Chen et al., 1995). Shelly fossils are abundant in the Arenig and Llanvirn formations. Most of those regional stratigraphical works and systematic publications on Early and Middle Ordovician brachiopods of South China employed the traditional British subdivision containing Tremadoc, Arenig and Llanvirn (Fortey et al., 2000). We subdivide these three stages into eight intervals (Figure 1), and all the discussions will be based on this temporal framework.
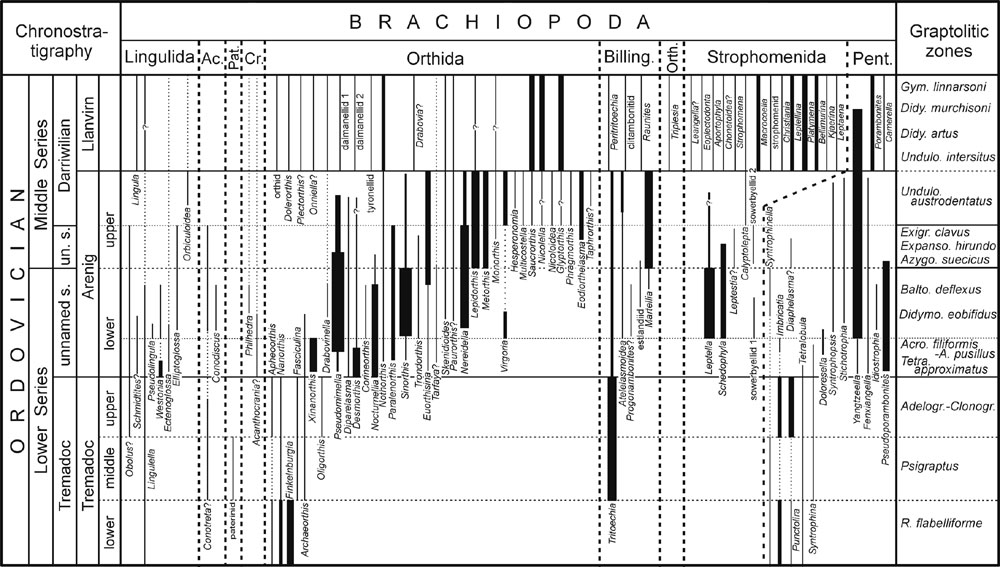
Figure 1. Stratigraphic range chart of Early to Middle Ordovician brachiopods of South China. Graptolitic zones are provided by Chen Xu and Zhang Yuandong. Ac., Acrotretida; Pat., Paterinida; Cr., Craniida; Billing., Billingsellida; Orth., Orthotetida; Pent., Pentamerida; un. s., unnamed stage.
1. Early Tremadoc (ET)
Only 8 genera were found belonging to 3 orders, 4 superfamilies and 6 families (Figure 1). Lingulella, Apheoorthis and Finkelnburgia were continuous from the Upper Cambrian while Nanorthis, Syntrophinella, Imbricatia, Punctolira and Tetralobula were new comers in South China. The occurrence of Nanorthis represents the first appearance of Orthoidea in South China. The Pentamerida, represented by four genera, is the third brachiopod order occupying the shallow water benthic regime herein.
2. Mid Tremadoc (MD)
Ten genera have been reported from South China within this interval (Figure 1). There are some major innovations on higher taxonomic levels. The number of brachiopod orders was doubled and three additional superfamilies (Acrotretoidea, Paterinoidea, and Polytoechioidea) appear. The Paterinida is a new comer represented by a single indeterminate genus that became disappeared from South China by the end of this interval. The most important change is the appearance of Tritoechia, a typical Ordovician cosmopolitan genus (Popov et al., 2001) that occurred at many horizons and localities in South China. It represents the first appearance of Billingsellida in South China.
3. Late Tremadoc (LT)
Although the number of higher taxa has slightly decreased (5 orders, 6 superfamilies and 9 families), there is an increase of generic number (Figure 1). Apheoorthis, Imbricatia and Punctolira disappeared during mid Tremadoc, and Obolus? and Oligorthis were new comers in South China. Oligorthis is limited within late Tremadoc. Within the Orthida, the familial and generic components are more than doubled (from 2 to 4 and 3 to 7).
4. Early early Arenig (EEA)
Altogether 25 genera belonging to 5 orders, 8 superfamilies and 14 families are known from various lithologies in this interval. The number of higher ranks seems stable. A major change happened to the Orthida that includes 4 superfamilies, 8 families and 11 genera, amongst which 10 genera are new comers in South China, and 5 families are represented by a single genus while 4 of them are new. Yangtzeella is widely distributed and very abundant on the Yangtze Platform in Arenig-Llanvirn rocks (Xu and Liu, 1984). It was also reported from the Arenigian rocks of Southeast Turkey (Cocks, 2000) and Northwest China (Xu and Sun, 1998). One of the most important innovations of this interval is the occurrence of Sinorthis and Euorthisina. Sinorthis, widely distributed in the Yangtze Region, usually acted as a dominant component of some shallow water benthic communities in South China. It was also reported from the Arenigian rocks of NE China (Liu et al., 1985), NW China (Xu and Sun, 1998) and France (Melou, 1982). Euorthisina is known to be widely distributed in China, Bolivia, Argentina and Bohemia (Xu et al., 1974; Havlícek and Branisa, 1980; Liu et al., 1985; Benedetto, 1998). Its appearance along with other small brachiopods represents the first successful attempt of brachiopods moving into deeper water benthic regime in South China.
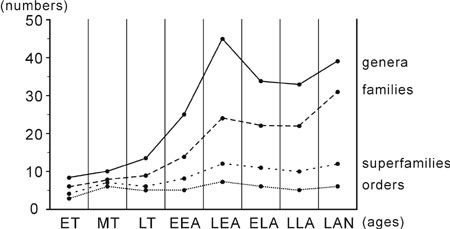
Figure 2. The change of brachiopod biodiversity of South China from early Tremadoc to Llanvirn on generic, familial, superfamilial and order levels. ET, early Tremadoc; MT, middle Tremadoc; LT, late Tremadoc; EEA, early early Arenig; LEA, late early Arenig; ELA, early late Arenig; LLA, late late Arenig; LAN, Llanvirn.
5. Late early Arenig (LEA)
The Ordovician radiation of brachiopods in South China reaches its first peak in this interval (Figure 2); it was mainly manifested by the generic and familial sharp increase: 45 genera belonging to 7 orders, 12 superfamilies and 24 families. The most important event in this interval is the first appearance of punctate orthoids (e.g., Nereidella) and pseudopunctate plectambonitoids (e.g., Leptella) in China. The Orthida was more developed than before having 5 superfamilies, 11 families and 18 genera (40% of the total). Both Sinorthis and Euorthisina are more abundant together with more associates constituting communities flourishing in shallow and deeper water regime. There occur a few genera (e.g., Pseudomimella and Fengxiangella) endemic to South China, and those cosmopolitans take more than 60%.
6. Early late Arenig (ELA)
After the first acme of Ordovician radiation, the brachiopod biodiversity decreases a little with a low generic recruiting rate (21%) and a comparatively high disappearing rate (38%) (Figure 3). According to this work, there occur 34 genera belonging to 6 orders, 11 superfamilies and 22 families. The Orthida is a dominant group having 5 superfamilies and 12 families, amongst which 8 are represented by a single genus. Metorthis and Eodiorthelasma are endemic to South China. Metorthis and its association took the predominant position of Sinorthis in the shallow water benthic regime through community replacement.
7. Late late Arenig (LLA)
The number of brachiopods in this interval is close to that in early late Arenig: 5 orders, 10 superfamilies, 22 families and 33 genera, but the generic and familial recruiting and disappearing rates are relatively high: (30%, 73%) and (18%, 36%), which means that faunal exchanges are significantly strengthened. The Orthida is still the most abundant including 5 superfamilies, 15 families and 23 genera (70% of the total), and is the only group that increases its diversity, whereas all the other groups decrease on both generic and familial levels. One of the most important events is the first occurrence of strophomenoids with a bilobed cardinal process in South China. The earliest known strophomenoid was reported from the Sandaokan Formation (early late Arenig) of NW China (Rong et al., 1999), and some representatives are found from the upper Daguanshan Formation (=Undulograptus austrodentatus Zone) in Shuanghe, Changning, southern Sichuan. Saucrorthis Xu and others (1974), a typical regional orthoid, was first named from the Shihtzupu Formation (late Llanvirn) of Zunyi, Guizhou and is known to occur from the lower Dashaba Formation (late Arenig) that might indicate its origination.
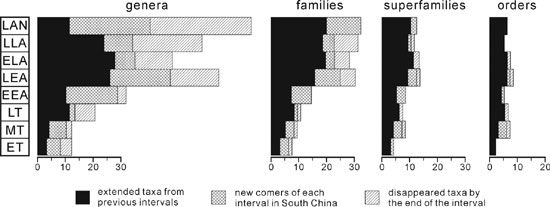
Figure 3. Diagrams expressing the macroevolution of Early to Middle Ordovician brachiopods of South China on different levels (genus, family, superfamily and order). Abbreviations see Figure 2.
8. Llanvirn (LAN)
After a period of depression, the brachiopods started to increase again by high generic recruiting and disappearing rates (72% and 90% respectively) representing the commencement of the second phase of the Ordovician radiation. Altogether 40 genera are known belonging to 6 orders, 12 superfamilies and 31 families. The Orthida is still an important group represented by 5 superfamilies, 12 families and 16 genera. The Strophomenida has greatly increased its biodiversity, from 1 superfamily to 2, 2 families to 10, and 2 genera to 14 (13 new comers). Particularly, the strophomenoids with bilobed cardinal process occurred at many localities with a high biodiversity and abundance. Triplesia from the Kuniutan Formation (Llanvirn) represents the first appearance of Triplesiidina in South China. Paleoecologically, the brachiopods of this interval are included into benthic communities or associations from BA 2 to BA 4.
Discussions and Conclusions
Early to Middle Ordovician brachiopods in South China show an overall evolutionary trend: more diversified since early Tremadoc with the change of taxanomic compositions of different groups and the expansion of benthic ecospace. The first biodiversity acme of the Ordovician radiation appeared in early to late early Arenigian transition, similar to the situation of Precordillera Basin where "the highest diversity occurred in the middle Arenig" (Waisfeld et al., 1999), but about four graptolitic zones earlier than the international trend (Sepkoski, 1995; Miller and Mao, 1998; Webby, 2000).
Systematically, the number of genera increased by nearly 6 times when the radiation got its first peak (from 8 in early Tremadoc to 45 in late early Arenig) and the number of family was quadrupled (from 6 to 24). The major taxonomic change occurred from early to late early Arenig. Those genera flourishing in Cambrian diminished since early Tremadoc, while more components of the Ordovician faunas, e.g., orthoids, clitambonitoids and porambonitoids, occurred and expanded regionally and even worldwidely.
Paleoecologically, those Cambrian brachiopods mainly lived in the nearshore shallow water benthic regimes (e.g., intertidal and upper subtidal) (Cocks, 1996). During the Tremadoc, most of the Yangtze Platform was covered by shallow water carbonates in which there are brachiopods of low diversity and abundance with impunctate and small shells, such as orthoids, clitambonitoids and porambonitoids. In the early Arenig, the vast area of the Upper Yangtze Platform was occupied by the typical shallow water brachiopod-dominated Sinorthis-Leptella Community. Up to late Early Ordovician (late early Arenig), some brachiopods (represented by Euorthisina together with Diparelasma, Nothorthis, Nocturnellia, Nereidella etc.) moved into a deeper water benthic regime (e.g., lower Meitan Formation at Shatuo, Yanhe, northeastern Guizhou) to form a unique community (Euorthisina Community) that is characterized by relatively high species diversity and small shell sizes including mixed brachiopod components: those shallow water constituents (e.g., Sinorthis, Leptella, Yangtzeella) being mainly euryplastic and those limited to deeper water (e.g., Euorthisina, Nereidella, Nocturnellia, Skenidioides) being usually stenotropic. This observation firmly supports Sepkoski and Sheehan (1983)’s statement that the Ordovician radiation began in onshore habitats and moved progressively to offshore environments.
Acknowledgements
This work has been supported by MST (G2000077700 and DEA20020-4), NSF (Nº 40002001) and CAS (KZCX2-SW-129) of China. We thank Zhang Yuan-dong for identifying graptolites and Chen Xu, Zhou Zhi-yi, Wang Yi, Zhang Yuan-dong, Fan Juan-xuan, Liu Jian-bo, Cheng Jin-hui and Xu Hong-he for their kindest help in the field. We also thank Profs. Guillermo Albanesi, Chen Xu, David Harper and Jisuo Jin for enlightening discussions and constructive suggestions on the manuscript.
References
Benedetto, J.L. 1998. Early Ordovician (Arenig) brachiopods from the Acoite and Sepulturas Formations, Cordillera Oriental, northwestern Argentina. Geologica et Palaeontologica, 32(1): 7-27.
Chen Xu, Rong Jia-yu, Wang Xiao-feng, Wang Zhi-hao, Zhang Yuan-dong and Zhan Ren-bin. 1995. Correlation of the Ordovician Rocks of China-Charts and Explanatory Notes. International Union of Geological Sciences, Publication, 31: 1-104.
Cocks, L.R.M. 1996. Lower Palaeozoic brachiopod communities. In: Copper, P. and Jin, J. (Eds.), Brachiopods-Proceedings of the Third International Brachiopod Congress. A. A. Balkema, Rotterdam: 69-71.
Cocks, L.R.M. 2000. The Early Palaeozoic geography of Europe. Journal of the Geological Society, London, 157: 1-10.
Cope, J.C.W. and Babin, C. 1999. Diversification of bivalves in the Ordovician. Geobios, 32(2): 175-185.
Droser, M.L. and Sheehan, P.M. 1995. Paleoecological significance of the Ordovician radiation and end Ordovician extinction: evidence from the Great Basin. In: Cooper, J.D. (ed.), Ordovician of the Great Basin. Pacific Section SEPM, Fullerton, California: 64-106.
Fortey, R.A., Harper, D.A.T., Ingham, J.K., Owen, A.W., Parkes, M.A., Rushton, A.W.A. and Woodcock, N.H. 2000. A revised correlation of Ordovician rocks in the British Isles. Geological Society Special Report, 24: 1-83.
Havlíèek, V. and Branisa, L. 1980. Ordovician brachiopods of Bolivia (Succession of assemblages, climate control, affinity to Anglo-French and Bohemian provinces). Rozpravy Èeskoslovenské Akademie vìd, Praha, 90: 1-54.
Liu Di-yong, Zhu Ci-ying and Xue Chun-ting. 1985. Ordovician brachiopods from northwestern Xiao Hinggan Ling, Northeast China. Bulletin of Shenyang Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences, 11: 1-46 (in Chinese with English abstract).
Melou, M. 1982. Brachiopodes articules. In: Babin, C. et al., Brachiopodes (Articules) et Mollusques (Bivalves, Rostroconches, Monoplacophores, Gastropodes) de l’Ordovicien inferieur (Tremaocien-Arenigien) de la Montagne Noire (France Meridionale). Societ d’Etudes Scientifiques de l’Aude, Memoire, 23-35.
Miller, A.I. and Mao, Shuguang. 1998. Scales of diversification and the Ordovician radiation. In: McKinney, M.L. and Drake, J.A. (Eds.), Biodiversity Dynamics-Turnovers of Populations, Taxa, and Communities. Columbia University Press, New York: 288-310.
Popov, L.E., Vinn, O. and Nikitina, O.I. 2001. Brachiopods of the redefined family Tritoechiidae from the Ordovician of Kazakhstan and South Urals. Geobios, 34(2): 131-155.
Rong Jia-yu and Chen Xu. 1987. Faunal differentiation, biofacies and lithofacies patterns of the Late Ordovician (Ashgillian) in South China. Acta Palaeontologica Sinica, 26(5): 507-535 (in Chinese with English summary).
Rong Jia-yu, Zhan Ren-bin and Xu Han-kui. 1999. The oldest known strophomenoid from the Sandaokan Formation (late Arenig, Ordovician) of Inner Mongolia, North China. Acta Palaeontologica Sinica, 38(1): 31-45.
Sepkoski, J.J. Jr. 1995. The Ordovician radiations: diversification and extinction shown by global genus-level taxonomic data. In: Cooper, J.D., Droser, M.L. and Finney, S.C. (Eds.), Ordovician Odyssey: Short Papers for the Seventh International Symposium on the Ordovician System. Pacific Section SEPM, Fullerton, California: 393-396.
Sepkoski, J.J. Jr. and Sheehan, P.M. 1983. Diversification, faunal change, and community replacement during the Ordovician radiations. In: Tevesz, M.J.S. and McCall, P.L. (Eds.), Biotic Interactions in Recent and Fossil Benthic Communities. Plenum Press, New York: 673-718.
Waisfeld, B.G., Sanchez, T.M. and Carrera, M.G. 1999. Biodiversification patterns in the Early Ordovician of Argentina. Palaios, 14: 198-214.
Webby, B.D. 2000. In search of triggering mechanisms for the great Ordovician biodiversification event. Palaeontology Down Under 2000, Geological Society of Australia, Abstracts 61: 129-130.
Xu Han-kui and Liu Di-yong. 1984. Late Lower Ordovician brachiopods of Southwestern China. Bulletin of Nanjing Institute of Geology and Palaeontology, Academia Sinica, 8: 147-235 (in Chinese with English summary).
Xu Han-kui, Rong Jia-yu and Liu Di-yong. 1974. Ordovician brachiopods. In: Nanjing Institute of Geology and Palaeontology, Academia Sinica (ed.), Handbook of Stratigraphy and Palaeontology in Southwest China. Science Press, Beijing: 144-154 (in Chinese).
Xu Han-kui and Sun Dong-li. 1998. Discovery of Early Ordovician brachiopods of Yangtze type in Karakorum Region and its significance. In: Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (ed.), Palaeontology of the Karakorum-Kunlun Mountains. Nanjing University Press, Nanjing: 145-162 (in Chinese with English summary).
Received: February 15, 2003
Accepted: June 15, 2003