Taxonomic turnover and community replacement in the Lower Ordovician limestones of the Argentine Precordillera
Marcelo G. Carrera1 and Norma Cech1
1 Cátedra de Estratigrafía y Geología Histórica Facultad de Ciencias Exactas Físicas y Naturales Universidad Nacional de Córdoba. Av. Velez Sársfield 299 (5000) Córdoba, Argentina. E–mail: mcarrera@com.uncor.edu
Keywords: Paleoecology. Limestones. Ordovician. Precordillera. Argentina.
Introduction
The fossiliferous Early Ordovician San Juan Formation in the Argentine Precordillera has been subject of detailed paleoecological studies in recent years (Carrera, 2001; Cech and Carrera 2002; Sánchez, et al., 2002 and references therein). These analyses are based on remarkable extensive taxonomic assessments that have been completed in the last twenty years.
Brachiopod biozones within the San Juan Formation (Benedetto, 2002) mark significant faunal turnovers, now well constrained by detailed conodont biostratigraphy (Albanesi et al., 1998, 2003; Lehnert, 1995). All these data, including information from other fossil groups, allow the establishment of 8 biotic intervals in the Ordovician of the Precordillera basin (Sánchez et al., 2002), three of which are included in the lower Ordovician limestones.
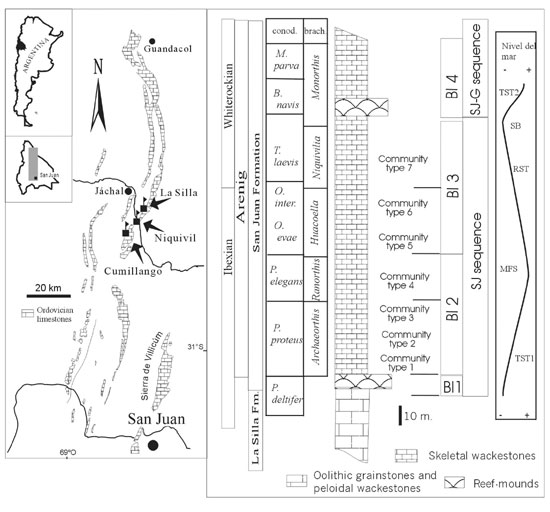
Cech and Carrera (2002) studied in detail the community replacement and distribution throughout the San Juan Formation in the Niquivil section (Figure 1). Updated paleoecologic information is included here from the adjacent more complete sections at La Silla and Cumillango Ranges, where different community types are established. Dominant taxa have been determined using biovolume values, a measure for biomass content in paleocommunities.
Paleoecologic analysis of global extinctions indicate that the relative magnitudes of changes measured by taxonomic diversity were not the same as the relative magnitudes of associated ecological changes (Bottjer et al., 2000; Droser et al., 2000). The aim of this preliminary contribution is to analyze and compare faunal turnovers based on taxonomic data and paleoecological changes occurring at community level in the Arenig limestones of the San Juan Formation.
This work was supported by grants from ANCYPT, PICT 5387 and CONICET, PIP 05/1316.
Paleoecology
Ordovician rocks in the Argentine Precordillera (Figure 1) are represented by Tremadocian carbonates (La Silla Formation) deposited in an isolated rimmed platform (Cañas, 1999) and muddy fossiliferous carbonates with open marine fauna (San Juan Fomation).
Figure 1. Location map and composite stratigraphic chart of the estudied sections, including community–type distribution. BI: Biotic intervals from Sánchez et al. (2002), sequence stratigraphy from (Cañas, 1999), brachiopod biozones from Benedetto (2002), condont biozones from Albanesi et al. (1998, and this volume).
Fossils assemblages in the La Silla Formation are very scarce with very few individuals recovered. In the Late Cambrian interval, the Plethopeltis trilobite biofacies has been recognized (Vaccari, 1995). During the Tremadocian, scarce remains of unidentifiable trilobites, nautiloids, gastropods and pelmatozoan ossicles occur, and laminar to domal thrombolites represent important components (Cañas, 1999).
In the San Juan Formation, level bottom communities are clearly dominated by suspensive–feeders, mainly brachiopods, sponges, echinoderms, and gastropods. Algae and microbial communities, including calcareous cyanobacteria, are represented by Girvanella, Nuia, and the problematical Calathium, and Fisherites.
The establishment of sponge–microbial reef–mounds (Cañas 1999, Carrera, 2001) at the base of the San Juan Formation (P. deltifer biozone) represents the onset of a transgressive event, the San Juan depositional sequence. This sequence boundary marks the most important faunal turnover in the basin for most of the components of the subsequent Arenigian communities belong to the Paleozoic evolutionary fauna. This limit is also the base for the first biotic interval of Sánchez et al. (2002) that includes the sponge–microbe reef–mounds.
Analysis of the community replacement is restricted to biotic intervals 2 and 3 which are included within the same depositional sequence. They record relatively similar environments, with a minor deepening from an inner to middle carbonate ramp (Cañas, 1999) during the O. evae biozone (Figure 1). Division of intervals 2 and 3 was based on taxonomic vertical distribution using holdover and carryover metrics (Sánchez et al., 2002).
Three particular boundaries are considered here: Archaeorthis–Ranorthis biozones boundary (within biotic interval 2), Ranorthis–Huacoella boundary (Intervals 2–3 limit) and Huacoella–Niquivilia boundary (within biotic interval 3, base of T. laevis, Lower–Middle Ordovician boundary).
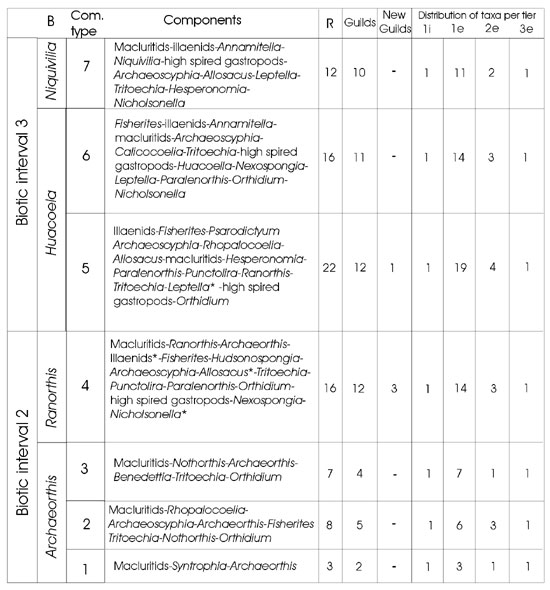
Table 1. Community types and distribution of guilds and tiers along the studied intervals. Community components ordering according dominance values. Crinoids fragments and fossil traces, not included in biovolume census, are considered in epifaunal and infaunal tiers. *: New guilds, R : richness, B: brachiopod biozones.
The Archaeorthis–Ranorthis boundary occurs within biotic interval 2 where low values of taxonomic turnover comprise the disappearance of three brachiopod genera (Sánchez et al. 2002). In paleoecological terms, it represents a replacement of community types. Community type 3 (table 1) is a group of successive local communities dominated by orthid brachiopods (Archaeorthis, Nothorthis) and gastropods. This community type is replaced by a more diversified community type 4, which is dominated by gastropods, orthid and pentamerid brachiopods and sponges. New guilds are added (Illaenus, Nicholsonella and Allosacus–Nexospongia) and a diversification within particular guilds occur.
The Ranorthis–Huacoella boundary is coincident with the boundary between biotic intervals 2 and 3 (BI2–BI3). Taxonomic turnover is mainly verified among trilobites genera, four of which disappeared and other four appeared. In addition, four new brachiopod genera are recorded in the interval 3 (Sánchez et al., 2002). The new community type 5, which records maximum richness, is dominated by trilobites and sponges. The reclining suspensive new guild is added, represented by the leptellinid brachiopods.
The Huacoella–Niquivilia boundary is also marked by a change in community type. Community type 6 dominated by receptaculitids, brachiopods (Tritoechia and Huacoella) and trilobites (Annamitella). Community type 7 is poorly diversified and dominated by gastropods, trilobites (Illaenus, Annamitella) and brachiopods (Niquivilia, Leptella). Although, the change in community type and the relatively important taxonomic turnover, neither guilds are added nor "filling–in" of previous guilds are recorded.
Discussion
Changes in community–types occur at all critical boundaries. New orthid–macluritacean community–type at the base of biotic interval 2 (BI2), and illaenid–receptaculitid–lithistid sponges occurred at Ranorthis–Huacoella boundary. These ecological changes imply a third level of ecological change (Bottjer et al., 2000; Droser et al., 2002).
Within biotic interval 2 (Archaeorthis–Ranorthis boundary) an important ecological change occurs. The ecological structure changed with more guilds recorded in the first and second level of tiering. In addition, three new guilds are recorded. However, this change is not associated with a taxonomic radiation, which seems to be delayed until the new guilds diversified at the next critical boundary.
According to taxonomic diversity, an important turnover occurs at the B.I.2–B.I.3 boundary. Ecological changes, in terms of tiering occupation and appearance of new guilds, are of comparably minor magnitude than those of the previous interval. The dominant biconvex brachiopods (orthids) of the epifauna–attached–suspension megaguild (Bambach, 1983) loses their dominance in community–type 5 and they are partially replaced by leptellinid brachiopods of the epifauna reclining suspension megaguild. The first appearance of this megaguild among brachiopods and the shifting in dominance from orthids to leptellinids represents a significant ecological change, and a third level filling of a megaguild (Bottjer et al. 2000, Droser et al. 2002). In terms of biomass, the contribution of brachiopods to the community structure is of minor significance. However, the installation of a new guild in an persistent ecological structure is an important feature, as well as its subsequent diversification in younger assemblages.
A decoupling of relative magnitudes of taxonomic turnover and ecological changes is difficult to sort out at this level of resolution. The minor taxonomic shift at the Archaeorthis–Ranorthis boundary is decoupled from the importance of ecological changes. At the biotic interval boundary, where a higher taxonomic turnover occurred, the ecological change is also important, although restricted to particular clades and influenced by the ecological features of the previous interval.
Differentiation and ordering of paleoecological levels at this resolution still needs further study. The real magnitude of the ecological signals (e.g., change of community type, appearance of new guilds) and their influence on the community structure must be reevaluated. Even so, much more detailed studies at community level are necessary for a better understanding of major paleoecological events.
References
Albanesi, G., Hunicken, M. y Barnes, C.R. 1998. Bioestratigrafía de conodontes de las secuencias ordovícicas del Cerro Potrerillo, Precordillera central de San Juan, R. Argentina. Actas de la Academia Nacional de Ciencias, Córdoba, 12: 7–72.
Albanesi, G. L., Carrera, M. G, Cañas F. L. and Saltzman, M. 2003. The Niquivil section, Precordillera of San Juan, Argentina, proposed GSSP for the Lower / Middle Ordovician boundary. (This volume)
Bambach, R.K. 1983. Ecospace utilization and guilds in marine communities through the Phanerozoic. In Tevesz M and Mc.Call., P. (Eds.), Biotic interactions in recent and fossil benthic communities Plenum press, New York: 719–746.
Benedetto, J.L. 2002. The Ordovician brachiopod faunas of Argentina: Chronology and biostratigraphic succession. In: Aceñolaza F.G. (ed.), Some aspects on the Ordovician of Argentina. INSUGEO, Serie de Correlación Geológica, Tucumán, 16: 87–105.
Bottjer, D. J., Droser, M., Sheehan, P. and McGhee Jr., G. 2000. The ecological architecture of major events in the Phanerozoic history of marine invertebrate life. In: W. Allmon and D. J. Bottjer (Eds.), Evolutionary Paleoecology. Columbia University Press: 35–61.
Cañas, F.L. 1999. Facies and sequences of the late Cambrian–early Ordovician carbonates of the Argentine Precordillera: A stratigraphic comparation with Laurentian platforms. In: Keppie D., and Ramos, V. (Eds.), Laurentia–Gondwana connections before Pangea, Geological Society of America, Special Paper 336: 43–62.
Carrera, M.G. 2001. Análisis de la distribución y composición de las biofacies de la Formación San Juan (Ordovícico temprano), Precordillera Argentina. Ameghiniana, 38 (2): 169–184.
Cech N. and Carrera, M.G. 2002. Dinámica de las comunidades arenigianas de la Formación San Juan (Ordovícico), Precordillera Argentina. Ameghiniana, 39 (1): 21–40.
Droser, M., Bottjer, D., Sheehan, P. and McGhee Jr., G. 2000. Decoupling of taxonomic and ecologic severity of Phanerozoic marine mass extinctions. Geology, 28(8): 675–678.
Lehnert, O. 1995. Ordovizische Conodonten aus der Präkordillere Westargentiniens: Ihre Bedeutung für Stratigraphie und Paläogeographie. Erlanger Geologische Abhandlungen, 125: 1-193.
Sánchez, T.M., Carrera, M. and Waisfeld, B. 2002. Hierarchical factors controlling faunal distribution: A case study from the Ordovician of the Argentine Precordillera. Palaios, 17: 309–326
Vaccari, N.E. 1995. Early Ordovician trilobite biogeography of Precordillera and Famatina, western Argentina: preliminary results. In: Cooper, J., Droser, M. L. and Finney, S. C. (Eds.), Seventh International Symposium on the Ordovician System, The Pacific section Society for Sedimentary Geology (SEPM), Fullerton, Book 77: 193–196.
Received: February 15, 2003
Accepted: June 15, 2003