Moulting in the Ordovician dalmanitid and acastid trilobites of the Prague Basin (Czech Republic)
Petr budil1 and Jana Bruthansova2
1 Czech Geological Survey, Klárov 3, Praha 1, 118 21, Czech Republic. E–mail: budil@cgu.cz
2 Palaeontological Department, National Museum, Václavské nám. 68, Praha 1, 115 79, Czech Republic. E–mail: jana.bruthansova@nm.cz
Key words: Dalmanitidae. Acastidae. Ordovician. Prague Basin. Czech Republic.
Introduction
The dalmanitid and acastid trilobites are among the most common Bohemian Ordovician trilobites. They have a wide stratigraphical range from the Lower Ordovician Klabava Formation to the Upper Ordovician Kosov Formation where they constitute the most important elements of the "Hirnantia fauna". The aims of this contribution are to evaluate the possible dalmanitid and acastid exuviae and to distinguish the main types of moulting positions of these trilobites, their occurrence in dependence on the phyllogenetical position and overall body morphology.
Geological setting
The Prague Basin originated during the extensive lower – middle Tremadocian transgression. It affected a new, narrow graben–like sedimentary space with its longitudinal axis along a subsiding SW–NE tectonic zone crossing to the former Príbram–Jince Basin (Havlícek in Chlupác et al., 1998) of Cambrian age. Both basins have been traditionally included into the classical Barrandian area that has been intensively studied for more than 200 years. During the Ordovician, the Prague Basin was filled mostly by siliciclastics and volcaniclastics of marine origin that were replaced by mostly carbonate sedimentation in the Silurian. Marine, mostly carbonate sedimentation persisted in the Prague Basin without any substantial break up to the Middle Devonian.
Types of the moulting configurations
In the Bohemian Ordovician dalmanitid and acastid trilobites, the moulting positions are very variable, especially in comparison with their relatives – phacopid trilobites (see Chlupác, 1977). Very often, even at one species, several modes of moulting occur together. Especially in the phyllogenetically earlier representatives (subfamily Zeliszkellinae Delo, 1935) the moulting variability is very high. The main observed types of the supposed moulting positions are shown in Figure 2.
Figure 1. The geographical position of the Prague Basin, Czech Republic (after O. Fatka, unpublished).
1) The most common mode of moulting is the separation of the cephalon from the complete thorax (see Figure 2.2), but without inversion typical for the "Salter´s position"(Speyer 1985). In some cases thoracopygon can be slightly overlapped by the cephalon (see Busch and Schwartz, 1985, a.o.). Ecdysial gape suture lies between cephalon and thoracopygon. Often (see Figure 2.4), the thorax is partially enrolled (see also Speyer (1985) at Greenops boothi).
2) On the contrary, the configurations similar to the true Salter´s position (see Richter, 1937; Chlupác, 1977; Speyer, 1985) are extremely rare, but observed in almost all species (see Figure 2.3).
3) Especially in Ormathops, but also in some other genera (Zeliszkella, Dalmanitina), the ecdysial gape suture is not placed exactly between the thorax and cephalon (cf. Speyer, 1985, Wenndorf, 1990). Frequently, with cephalon, at least two or three segments are still articulated and this part of the exoskeleton is separated from the articulated rest of the thorax + pygidium (see Figure 2.5).
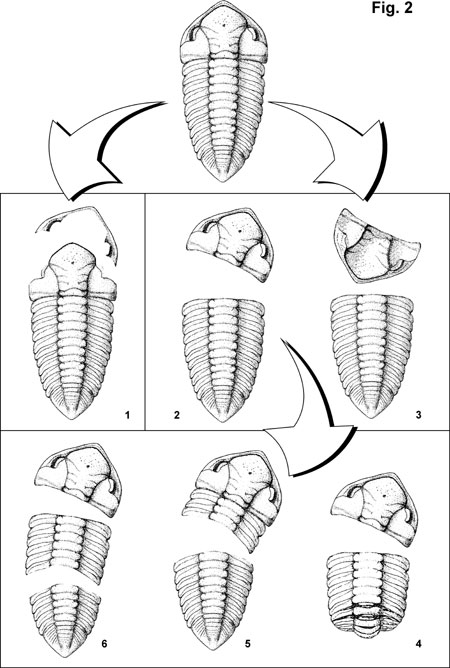
Figure 2. The main observed types of the moulting positions at Bohemian Ordovician dalmanitid and acastid trilobites.
4) In other cases, a thorax is crumbled into the two or more independent parts. A similar moulting strategy in Shumardia (Conoprys) salopiensis was interpreted by Fortey and Owens (1991) as escapist, e.g., used if all efforts of the animal to remove old exoskeleton were not successful. Similar mode of moulting was described by Snajdr (1990) in the Bohemian paradoxiid trilobites. In Ormathops atavus (Barrande), an incomplete cephalothoraxex with preserved hypostome in situ were rarely found. If they are exuviae, it could be considered as an extreme case of articulation of the hypostoma in this species.
The function of genal suture was probably preserved at least in some cases in Ormathops atavus and Ormathops barroisi (Kloucek). In a few complete adult samples librigenae are completely missing (see Figure 2.1), so that this configuration partially resembles the axial shield (Henningsmoen, 1973). Positions similar to the classical Salter’s position with the librigenae separated from the cranidium were studied in Ormathops atavus (Barrande).
Separated free cheeks have been also observed at isolated cephala (1–2%) in Dalmanitina cilinensis Snajdr, D. proaeva proaeva (Barrande), D. proaeva elfrida Snajdr, Eudolatites dubius (Barrande) and Duftonia morrisiana (Barrande). In Dalmanitina cilinensis, isolated and broken free cheeks are relatively common in the organic detritus of the lower Berounian Revnice quartzite. In that case, without any other supplementary features supporting that separation of librigenae occured as a result of moulting, also post–ecdysial and/or post–mortal disintegration of the exoskeleton and subsequent transport of the organic remains by sea currents should be also considered.
The lack of pygidium in complete cephalothoraxes has been observed but it is not clear if it is really a moulting position as well as like in enrolled samples without pygidium.
On the contrary, the configurations described by McNamara and Rudkin (1984) in pterygometopid trilobite Toxochasmops extensus (isolated cephala and pygidia lying close each other, often with slight imbrication, without, or with few thoracic segments only) were not observed at the Bohemian material.
Conclusions
A wide variability of the moulting positions was observed at Bohemian Ordovician dalmanitid and acastid trilobites, especially in the phylogenetically early representatives. In the advanced dalmanitids and in most of acastids, few types of moulting positions (type 1, 2, 3) prevail and diversity markedly decreased during the Upper Ordovician. No important differences in the prevailing types of moulting were observed in dalmanitids and early acastids that occur together in the Prague Basin. The diversity and prevailing types of the moulting mode seems to be rather the function of the overall body morphology than the virtual phyllogenetical relations of the species. For example, a morphologically similar, but probably non–relative Ormathops atavus (Barrande) and Phacopidina quadrata (Hawle et Corda) have very similar percentage occurrence of the main moulting types. On the contrary, even in very closely related species of the same genus (Ormathops, Zeliszkella), surprising differences in the variability of moulting were observed.
Acknowledgements
This study was conducted with the financial support of the Grant Agency of the Academy of Sciences of the Czech Republic project Nº B 3407201.
References
Busch, R. M. – Swartz, F. M. 1985. Molting and description of a new homalonotid trilobite from Pennsylvania. Journal of Paleontology, 59: 5, 1062–1074.
Chlupác, I. 1977. The phacopid trilobites of the Silurian and Devonian of Czechoslovakia. Rozpravy Ústredního ústavu geologického, 43: 1–172.
Chlupác, I. – Havlícek, V. – Kríz, J. – Kukal, Z. – Storch, P. 1998. Palaeozoic of the Barrandian (Cambrian to Devonian). Czech Geological Survey, Prague.
Fortey, R. – Owens, R. 1991. A trilobite fauna from the highest Shineton Shales in Shropshire, and the correlation of the latest Tremadoc. –Geological Magazine, 128, 437–464.
Henningsmoen, G. 1973. Moulting in trilobites. – Fossils and Strata, 4, 179–200. Oslo.
McNamara, K, J. and Rudkin, D. M. 1984. Techniques of trilobite exuviation. – Lethaia, 17, pp. 153–173. Oslo.
Richter, R. 1937. Vom Bau und Leben der Trilobiten. 8. Die "Salterische Einbettung" als Folge und Kennzeichen des Häutungs–Vorgangs. Senckenbergiana 19: 413–431.
Snajdr, M. 1990. Bohemian trilobites. 265 pp., Czech Geological Survey, Praha.
Speyer, E. 1985. Moulting in phacopid trilobites. Transactions of the Royal Society of Edinburgh: 76, 239–253.
Wenndorf, K. W. 1990. Homalonotinae (Trilobita) aus dem Rheinischen Unter–Devon. – Palaeontographica (Abt. A) 211, 1–184
Received: February 15, 2003
Accepted: June 15, 2003