The conodont succession from the proposed Gssp for the Middle Ordovician base at Huanghuachang Section, Yichang, China
Wang Xiaofeng1, Li Zhihong1, Chen Xiaohong1 and Wang Chuanshang1
1 Center for Stratigraphy and Palaeontology, Yichang Institute of Geology and Mineral Resources,Yichang, Hubei 443003, China. E–mail: wangxf@public.yc.hb.cn
Key words: Conodonts. Stratotype. Lower/Middle Ordovician boundary. Huanghuachang. China.
Introduction
The Huanghuachang section was proposed as the GSSP for the base of the Middle Ordovician (Wang et al., 2002). The conodont succession across the boundary interval is well developed in the Lower Dawan Formation with a great abundance and diversity (Zeng et al., 1983; An, 1987; Ni, 1987 in Wang et al., 1987; Wang Z. et al., 1996). Both the warmer water (Midcontinental Realm) and the cool water (the N. Atlantic Realm) conodonts are all found in the boundary interval. The present discussion is based largely on collection and process during last two years.
Conodont zonation and correlation
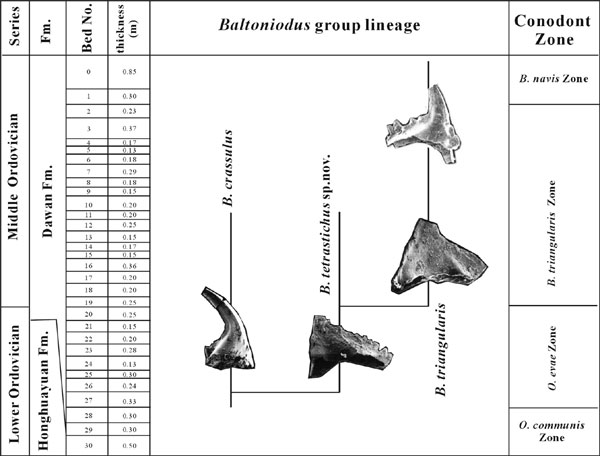
Recent detailed study further identifies a complete and well–known conodont succession can be recognized from the uppermost Honghuayuan Formation to the Lower Dawan Formation. The Oepikodus communis Biozone with 1.5m thick covers the uppermost Honghuayuan Formation to the lowest Dawan Formation (Hoh31– Hod 28). Its base and top are separately defined by the FAD of the index species and Oepikodus evae (Plate 1, Figures 18, 23–24). The latter first appears within a thick–bedded glauconitic bioclastic limestone (Hod 27), 0.6 m above the base of the Dawan Formation and ranges upwards to the most of B. triangularis Biozone. Associated major conodonts include Baltoniodus crassulus, Acodus? russoi, Protopanderodus gradatus etc. in the lower part besides of Scolopodus rex , Oepikodus communis, Bergstroemognathus extensus from the underlying O. communis Biozone. B. tetrastichus Li sp. nov., Scolopodus praerex, Drepanoistodus forceps and Oepikodus intermedius occur in the middle and upper parts of the biozone. The B. triangularis Biozone is defined by the FAD of the index species (Figures 1–2, Plate 1, Figures 1–7) at base (Hod 19) and of B. navis at top (Hod 6). The base of the bioizone is located at 2.6m above the base of Dawan Formation and 0.2 m below the level with the FAD of T. laevis (Plate 1, Figures 8–12). Associated principal conodonts comprise B.triangularis, Tripodus laevis, Stolodus stola, Paltodus cf. subaequalis, Periodon flabellum and some elements ranging up from the underlying O evae Biozone. The level with FAD of Baltoniodus navis is situated at 6.58 m (Hod 1) above the base of Dawan Formation in the upper Lower Dawan Formation. Most conodonts of the biozone are coming from the underlying B. triangularis Biozone. Subsequent P. originalis Biozone covers the Middle Dawan Formation. Besides, there are minor Protoprioniodus aranda Cooper are recorded in the Lower B. navis Biozone at the section. This species,
Figure 1. Showing the evolving lineage of Baltoniodus group across the Lower/ Middle Ordovician boundary at Huanghuachang section (Based on Li Zhihong)
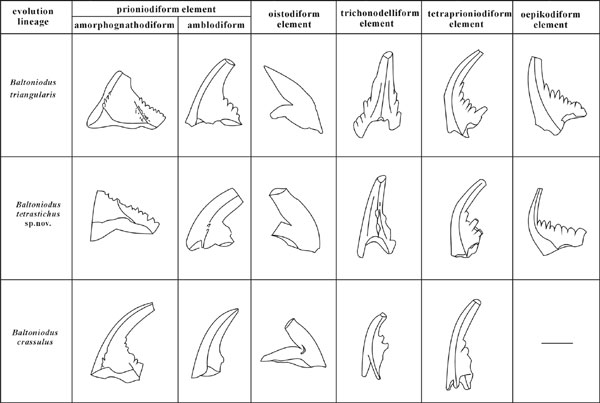
Figure 2. Showing the principal features of Baltoniodus tetrastichus Li sp. nov. and its evolving relationship with B. crassulus and B. triangularis.

Plate 1. Important conodonts collected from the Lower/Middle Ordovician Boundary interval (Lower Dawan Formation) at the Huanghuachang Section, identified by Li Zhihong. All specimens are deposited in the Museum of Yichang Institute of Geology and Mineral Resources (YIGMR).
1–7 Baltoniodus triangularis (Lindström, 1955)
1,2,5. amorphognathodiform element, lateral view, x120, x100, x80, Hod–9/2275, Hod–8/2283. 3.amblodiform element, lateral view, x120, Hod–9/2274. 4. trichonodelliform element, posterior view, ¡Á80, Hod–8/2269. 6. oepikodiform element, lateral view, x150, Hod–9/2278. 7. oistodiform element, lateral view, x180, Hod–8/2287.
8–12 Tripodus laevis Bradshaw, 1969
8.oistodiform element, lateral view, x300, Hod–2/5284. 9.drepauodontiform element, lateral view, x180, Hod–16/5320. 10. paltodiform element, lateral view, x230, Hod–16/5247. 11. trichonodelliform element, lateral view, x200, Hod–3/7048. 12. distacodiform element, lateral view, x120, Hod–17/7052.
14–16, 17a,17b, 25 Baltoniodus tetrastichus Li sp. nov.
14. amblodiform element, lateral view, x180, Hod–25/2327. 15. oistodiform element, lateral view, x120, Hod–16/2374. 16. tetraprioniodiform element, lateral view, x120, Hod–24/2472. 17a. amorphognathodiform element, x180, Hod–24/2623. 17b. oral view, x150, Hod–24/2324. 25. trichonodelliform element., posterior view, x180, Hod–25/ 2332.
13,19–22 Baltoniodus crassulus (Lindström, 1955)
13. amorphognathodiform element , lateral view, x130, Hod–21/5307. 19. prioniodiform element, lateral view, x120, Hod–13/2255. 20,21. ramiform element, lateral view, x150, Hod–16/2370, Hod–26/2316. 22. oistodiform element, lateral view, x150, Hod–26/2358.
18, 23–24 Oepikodus evae (Lindström, 1955)
18. oepikodiform element, lateral view, x100, Hod–25/ 2351. 23. oistodiform element, lateral view, x120, Hod–11/ 2266. 24. prioniodiform element, lateral view, x120, Hod–25/ 2336–1.
however, is found recently from the Upper O. evae Biozone to the Lower B. triangularis Biozone at the Wengxigou section of Chengkou, Chongqing. The conodonts succession identified from the Lower Dawan Formation has best potential for use as primary guide in the correlation and definition of the Lower/Middle Ordovician boundary. Comparing with T. laevis the FAD of Baltoniodus triangularis is chosen as the boundary biomark for the definition of the middle Ordovician base because the boundary between the O. evae and B. triangularis Biozones is widespread in the Yangtze platform, China (Wang et al., 1992) and Baltoscania (Löfgen, 1994, Bagnoli and Stouge, 1997). More importantly an excellent evolving lineage from Baltoniodus crassulus to B. triangularis via Bltoniodus tetrastichus Li sp. nov. (Figures 1–2, Pl. 1, Figures 14–16, 17a, 17b, 25) can be observed in the boundary interval. Baltoniodus crassulus (Figures 1–2, Pl. 1, figs.13, 19–22) was ever considered as the ancestor of B. triangularis (Klapper et al., 1977, p. 81). It first appears at the level (Hod 27), 0.6m above the base of the Dawan Formation along with O. evae and ranges upwards into the B. triangularis Biozone. Bltoniodus tetrastichus Li sp. nov., an intermediate new taxon between B. crassulus and B. triangularis (Figures 2–3), is characterized by the amorphognatodiform of multielement apparatus with 3 costae extending towards basally and forming an anterior process, a laterior process and a posterior process, with its second node sitting on the posterior process, bearing a costa extending downwards and backwards to the basal margin. It first appears at Hod 26, 0.3m above the FAD of B. crassulus, and gradually develops into B. triangularis with stretching of posterior–lateral carina, increasing of denticle number on the posterior process, and extending toward back and expending of basal cavity. Moreover, the appearance of T. laevis at 0.2 m above the base of B. triangularis Biozone, and associated chitinozoans, graptolites, acritarchs, brachiopods and trilobites in the boundary interval of the section, provide excellent ties for precise global correlations of the boundary in both shelly and shaly facies (Wang et al., 2003 in this volume).
Conclusions
Our study has further confirmed that the conodont succession from the Lower Dawan Formation at the Huanghuachang section is one of the most complete and well–represented sequences across the Lower and Middle Ordovician boundary interval in the world. The first appearance of Baltoniodus triangularis (Lindström, 1955), at 2.6m (the base of Hod 19) above the base of the Dawan Formation, is proposed as conodonts icon for the definition of the Middle Ordovician base and the first appearance of Tripodus laevis Bradshaw, 1969, at the level (Hod17), 0.2m above the boundary biozone at the proposed boundary stratotype, can be taken as a reasonable proxy for the boundary correlation. The proposed Middle/ Lower boundary can be recognized and correlated globally with high precision.
Acknowledgements
This research is financially supported by the NSFC, CCS and CGS. Special thanks are expressed to Stan Finney, Guillermo L. Albanesi, Stig Bergström, Svend Stouge and Wang Zhihao for their suggestions, comments and examinations of conodonts, respectively.
References
An T. X.1987. Paleozoic conodonts of southern China. Beijing: Science Press: 1–238.
Bagnoli G. and Stouge S. 1997. Lower Ordovician (Billingenian–Kunda) conodont zonation and provinces based on sections from Horns Udde, north Öland, Sweden. Bollettino della Società Paleontologica Italiana, 35 (2): 109–163, 8pls.
Bradshaw L.E. 1969. Conodonts from the Fort Pena Formation (Middle Ordovician), Marathon Basin, Texas. Journal of Paleontology, 43 (5): 1137–1168.
Chen X.H., Wang X.F., Li Z.H., Wang C.S., Zhang M. 2002. The chitinozoan assemblages across the Middle and Lower Ordovician boundary at the Huahuachang section , Yichang, Hubei, China and its stratigraphical significance. Journal of Stratigraphy, 26 (4): 241–247 (in Chinese with English abstract).
Lindström M. 1955. Conodonts from the lowermost Ordovician strata of South–central Sweden. Geologiska Foreningens I Stockhol Förhandlingar, 76 (4): 517–803.
Löfgren A. 1994. Arenig (Lower Ordovician) conodonts and biozonation in the eastern Siljan district, central Sweden. Journal of Paleontology, 68 (6): 1350–1368.
Wang X.F., Ni S.Z., Zeng Q.L., Xu K.H., Zhou T.M., Li Z.H., Xiang L.W. and Lai C.G. 1987. Biostratigraphy of the Yangtze Gorges area (2), Early Paleozoic. Geological Publishing House, Beijing: 1–641 (in Chinese with English summary).
Wang X.F., Chen X. and Erdtmann B. –D. 1992. Ordovician chronostratigraphy—A Chinese approach. In: Webby B.D. and Lauries J. R. (Eds.), Global Perspectives on Ordovician Geology, Ratterdam: 35–55.
Wang X.F., Chen X.H., Li Z.H., Wang C.S. 2002. The Huanghuachang section, near Yichang, China, proposed as global stratotype for the base of the Middle Ordovician Series. In: SOS, IUGS (Eds.) Internet web: 1–13.
Wang X.F., Chen X.H., Li Z.H., Wang C.S. 2003. The Huanghuachang section, potential as global stratotype for the base of Middle Ordovican Series. (This volume).
Wang Z.H., Bergström, S.M. and Lane, H.R. 1996. Conodont province and biostratigraphy in Ordovician of China, Acta Palaeontologica Sinica, 35 (3): 26–58.
Zeng Q.L., Ni S.Z., Xu K.H., Zhou T.M., Wang X.F., Li Z.H., Xiang L.W. and Lai C.G. 1983: Division and correlation of Ordovician System in East Yangtze Gorges Area. Bulletin of the Yichang Institute of Geology and Mineral Resources, CAGS, 6: 1–56.
Received: February 15, 2003
Accepted: June 15, 2003