Ordovician acritarchs from northwestern Argentina: new insights into the biostratigraphy and paleonvironmental aspects of the Central Andean Basin and Famatina
Claudia V. Rubinstein1
1 CONICET; Unidad de Paleopalinología, IANIGLA, CRICYT, 5500 Mendoza, Argentina.
Key words: Acritarchs. Biostratigraphy. Paleoenvironments. Paleogeography. Argentina.
Introduction
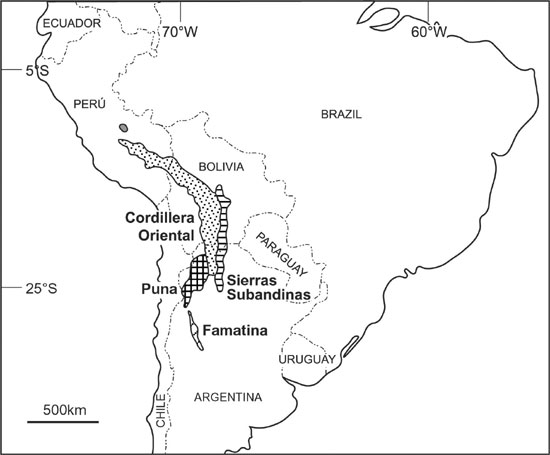
Ordovician sedimentary rocks are widely distributed in Argentina. The largest exposures correspond to the Central Andean Basin (Figure 1), comprising the Puna, Cordillera Oriental (Eastern Cordillera), and Sierras Subandinas (Subandean Ranges), where strata range in age from the Tremadoc to the Ashgill. Traditionally, the Puna, Cordillera Oriental and Sierras Subandinas have been considered as separate basins, especially because of their Cenozoic geological history, which has imprinted the present morphology. This interpretation was supported by paleontological, lithological and stratigraphical differences, although not always relevant enough to justify the separation of these basins during Ordovician time.
New investigations help explain these differences in the context of an integrated foreland basin model for the Ordovician of the Central Andean Region, where a continuous link of variable depositional settings (including inshore–offshore gradients) is represented (Astini and Marengo, 2003). In the light of this recent interpretation, an integrated reassessment of acritarch biostratigraphy (Figure 2) and paleonvironmental aspects is presented in which acritarch results are compared to those of the Famatina basin (Figure 1).
Central Andean Basin
In this new scenario, diversified acritarch assemblages from the Cordillera Oriental correspond to the platform facies of the foreland basin. Ordovician acritarch investigations began in the Cordillera Oriental, and for many years they have been sparse and have not allowed the elaboration of a regional biostratigraphic scheme. Pioneer studies involved Tremadoc and Arenig strata from the Cordillera Oriental (Volkheimer et. al., 1980; Bultynck and Martin 1982). These were followed by an integrated study of upper Tremadoc to middle Arenig acritarchs and graptolites from the Cordillera Oriental that has led to a precise biostratigraphy for the region, and has contributed to an assessment of its paleogeographical position (Rubinstein et al., 1999; Rubinstein and Toro, 2001). The richest acritarch assemblages come from the B. deflexus and D. bifidus graptolite biozones, where relevant biostratigraphical taxa such as Coryphidium bohemicum, Arbusculidum filamentosum and Aureotesta clathrata are present, thus indicating peri–Gondwana affinities (Brocke et al., 1995). Moreover, the presence of the messaoudensis–trifidum acritarch assemblage, characteristic of the Tremadoc–Arenig boundary in most peri–Gondwana areas (Servais and Molyneux, 1997), has been confirmed by the presence of Coryphidium, Cymatiogalea messaoudensis and C. deunffi in strata related to the A. murrayi graptolite Biozone (Rubinstein and Toro, 2002).
Figure 1. Location map.
Rubinstein et al. (in press) recently documented an upper Cambrian acritarch assemblage, probably close to the Cambrian–Ordovician boundary, in the Cordillera Oriental. This assemblage provides valuable information for a more accurate definition of the Cambrian–Ordovician boundary. Interestingly, the presence of acritarch taxa such as Saharidia and Vulcanisphaera africana which span the systemic boundary, have also been found in lower Tremadoc strata (Bultynck and Martin, 1982; Aráoz and Vergel, 2001).
Distal environmental settings of the foreland basin crop out in the Puna, where Ordovician–Silurian transitional strata have yielded a relatively well–preserved and diversified acritarch assemblage (Rubinstein and Vaccari, in press). Identification of the Ordovician–Silurian boundary on the basis of acritarchs is complicated because of the extinction caused by the Hirnantian glaciation. Many typical Ordovician acritarchs disappeared in the latest Ordovician, while those with Silurian characteristics first appeared in the Upper Ordovician (Molyneux et al., 1996, Martin, 1988).The Puna acritarch assemblages correspond to a shallow marine environment which proximity to land is suggested by the abundance of land–derived cryptospores. Acritarchs include Upper Ordovician species such as Eupoikilofusa striata and Villosacapsula cf. V. setosapellicula, that coexist with the typical Llandovery species Dactylofusa estillis. Based on lithological and palynological evidence, an early Llandovery, or even a late Hirnantian (post–glacial) age has been proposed for the bearing strata.
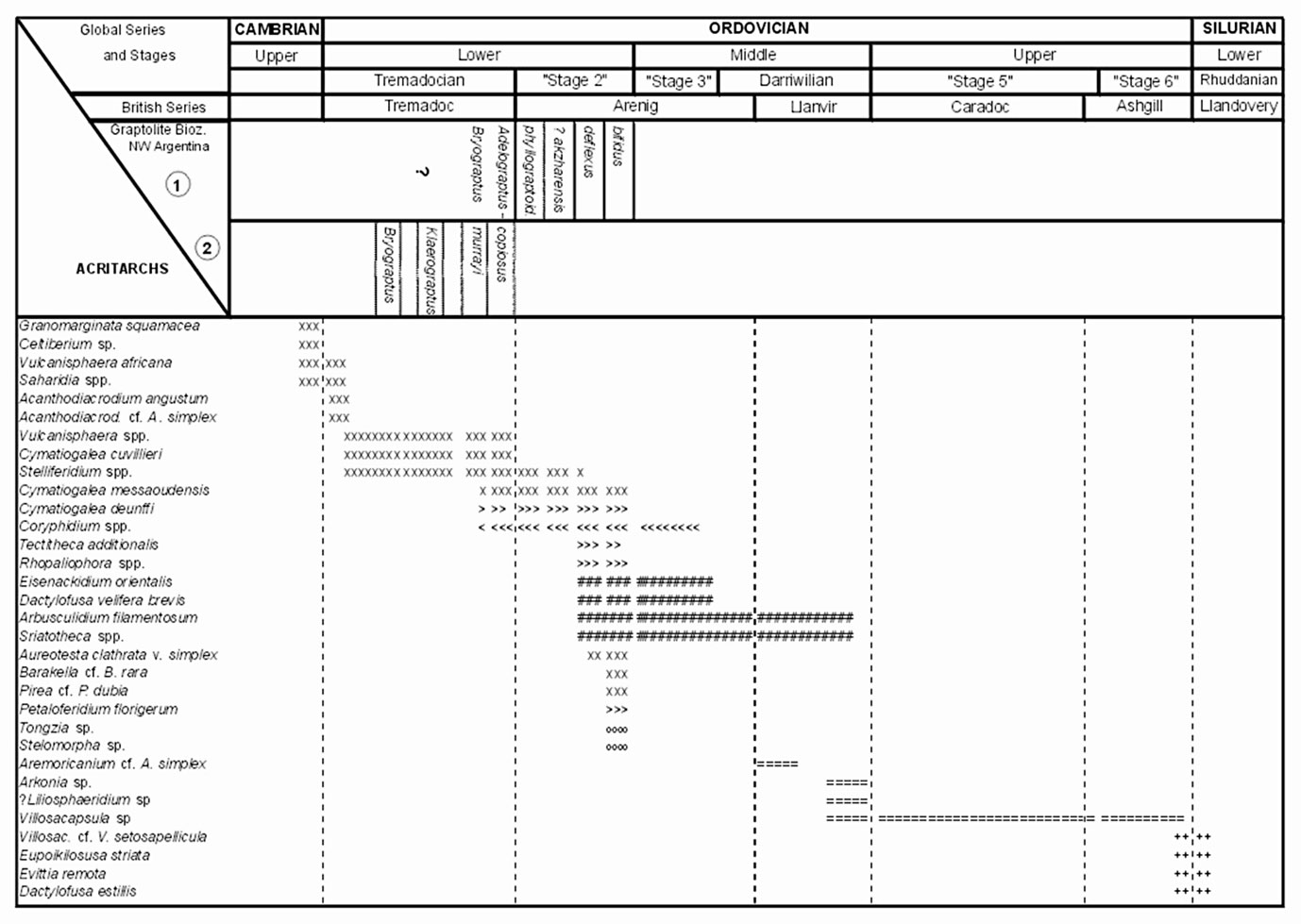
Figure 2. Distribution of selected acritarch taxa throughout the Ordovician of northwestern Argentina, and correlation with graptolite biozones. Graptolite Biozonations from the Cordillera Oriental: 1) Toro (1997), 2) Ortega and Albanesi (2002). Acritarch distribution: Cordillera Oriental (xxx): Aráoz and Vergel (2001), Ottone et al., (1992, 1995), Rubinstein (1997), Rubinstein and Toro (1999, 2001, 2002); Rubinstein et al., (1999) Rubinstein et al., (in press), Volkheimer et al., (1980). Famatina (ooo): Rubinstein and Astini (2000), Rubinstein (unpublished data). Puna (+++): Rubinstein and Vaccari (in press). Sierras Subandinas (===): Rubinstein, 2003. Acritarchs present in more than one basin: Cordillera Oriental and Famatina (>>>); Cordillera Oriental and Sierras Subandinas (<<<); Easten Cordillera, Sierras Subandinas and Famatina (###).
The proximal environmental facies of the Central Andean Basin are well documented in the Sierras Subandinas, where significant relative sea–level fluctuations are evident. This outermost region of the foreland basin is characterized by the alternation of shallow marine deltaic systems and estuarine environments (Astini and Marengo, 2003). Preliminary palynological results of Middle to Upper Ordovician deposits indicate a relationship between palynomorphs and depositional settings (Rubinstein, 2003).
The lowest Ordovician unit (Zanjón Formation) yielded an interesting acritarch assemblage containing Coryphidium sp., Striatotheca spp., Arbusculidium filamentosum and Eisenackidium orientalis. This assemblage enables correlation with acritarchs of the Acoite Formation (deflexus and bifidus graptolite biozones) from the Cordillera Oriental. The Zanjón Formation is considered middle to late Arenig in age, based on acritarchs and stratigraphical correlations.
The Labrado Formation is divided into two members. The lower Laja Morada Member, late Arenig in age, represents exhumation during relative sea level drop (Astini and Marengo, 2003). Consequently, the only palynomorphs present are leiospheres and algal cysts, neither of which has stratigraphic value. The upper Lagunillas Member, early to middle Llanvirn in age, yielded not only such acritarchs as Aremoricanium cf. A. simplex, but also leiospheres and probable pre–cryptospores, (sensu Richardson 1996) suggesting a proximal environment.
A maximum flooding surface constitutes the transition between restricted estuarine facies of the Lagunillas Member and the open marine deposits of the Capillas Formation. The latter was dated as late Llanvirn to basal Caradoc by means of Sacabambaspis janvieri (Vertebrata). This formation yielded a rich and well–preserved acritarch assemblage containing such taxa as Arbusculidium filamentosum, Striatotheca spp, Arkonia sp., Liliosphaeridium? and Villosacapsula sp.
The Centinela Formation, no younger than late Caradoc, contains mainly leiospheres, pre–cryptospores and simple acanthomorphitic acritarchs, in coincidence with a possible progradation of localized deltaic complexes. Upward, a regional unconformity correlate with a major eustatic sea–level drop, caused by the Hirnantian glaciation, separates the Centinela Formation from the uppermost Zapla Formation. The Zapla glacial horizon, probably Hirnantian in age, yielded acritarch assemblages dominated by Villosacapsula sp. and subordinate leiospheres and cryptospores (mainly tetrads).
Famatina
Several authors have interpreted the Ordovician of the Famatina Region as a subduction related active volcanic–arc located in the Gondwana margin during the approach of the Precordillera derived exotic terrane (Astini, 1999). Consequently, the Famatina was a complex volcanic terrane, such that sedimentary record was controlled by the interaction between vulcanism, tectonism and sea level changes. An upward shallowing continuous record, deposited in a marine environment, is identified in the Arenig sedimentary–volcanic succession of the Famatina., with increasing volcanic activity toward the top. Thus, the sedimentary and paleontological record is typical of island arc settings (Astini, 1999).
The Arenig sedimentary– volcanoclastic succession is represented by the Famatina Group, which includes the Suri and Molles formations. Acritarchs from both formations are middle Arenig in age and independently dated by conodonts from the upper part of the Oepikodus evae Zone (Rubinstein and Astini, 2000; Rubinstein 2001a, 2001b and unpublished data).
The Suri Formation shares several acritarch taxa with coeval deposits of the Cordillera Oriental and Sierras Subandinas. However, a notable increase in peteinoid acritarchs and representatives of Rhopaliophora is also noted. Significant taxa include Arbusculidium filamentosum, Striatotheca sp., Eisenackidium orientalis, Dactylofusa velifera forma brevis and Stelomorpha sp. Even though the mixture with middle to low latitude taxa, mainly with those of South China (Stelomorpha sp., abundance of peteinoid acritarchs and Rhopaliophora), acritarchs of the Suri Formation show clear peri–Gondwana affinities (Playford et al., 1995).
The panorama changes drastically in the Molles Formation. Characteristic elements of the cold water peri–Gondwana Realm completely disappear, and taxa such as Baltisphaeridium, Rhopaliophora, Tongzia, Peteinosphaeridiun, and other peteinoid forms, typical of intermediate to low latitudes related to South China, Australia and Baltica become predominant (Tongiorgi et al., 1995; Tongiorgi and Di Milia, 1999, Raevskaya, 1999). Changes in acritarch assemblages between the two units is considered a consequence of local conditions, produced by an extremely stressful volcanic–arc environment. This interpretation is also supported by conodont data (Albanesi and Astini, 2000).
Conclusions
Even at this preliminary stage of analysis, it is evident that acritarchs can provide a useful tool for biostratigrahy and correlation within the basins of northwestern Argentina. In addition, a relationship between acritarchs and depositional settings can be made.
Acknowledgements
Thanks to R. Wicander and G. Ottone for the review of the manuscript and useful suggestions. The research was supported by the CONICET, FONCYT, and Fundación Antorchas.
References
Albanesi, G. L. and Astini, R.A. 2000. New conodont fauna from Suri Formation (Early–middle Ordovician), Famatina System, western Argentina. Ameghiniana, 37(4), Suplemento, Resúmenes: 68.
Aráoz, L. and Vergel, M. d. M. 2001. Acritarcos del Ordovícico Inferior (Formación Santa Rosita) en el Abra de Zenta, Cordillera Oriental, Argentina. V Jornadas de Comunicaciones. Serie Monográfica y Didáctica. Nº 41. Facultad de Ciencias Naturales e I.M.L, Universidad Nacional de Tucumán: 55.
Astini, R. A. 1999. El Ordovícico del Sistema del Famatina. In: González Bonorino G., Omarini, R. and Viramonte, J. (Eds.). Geología del Noroeste Argentino. Relatorio XIV Gongreso Geológico Argentino, 1: 152–159.
Astini, R.A. and Marengo, L. 2003. Sequence Stratigraphy Of Perigondwanic Ordovician Clastics At The Outermost Reach Of The Protoandean Foreland, Sierras Subandinas, Argentina. 3º Latin American Congress of Sedimentology. Extended Abstracts Volume.
Brocke, R., Fatka, O., Molyneux, S. G. and Servais, T. 1995. First appearance of selected early Ordovician acritarch taxa from peri–Gondwana: Ordovician Odyssey Short papers for the Seventh International Symposium on the Ordovician System, SEPM 77: 473– 476.
Bultynck, P. and Martin, F. 1982. Conodontes et Acritarches de l’ Ordovicien Inférieur et acritarches du Silurien inférieur de la partie septentrionale de la Cordillère Argentine. Bulletin de l’ Institut royal des Sciences naturelles de Belgique, Sciences de la Terre. 53 (4): 1–21.
Martin, F. 1988. Late Ordovician and early Silurian acritarchs. Bulletin of the British Museum of Natural History (Geology), 43: 299–309.
Molyneux, S. G., Le Hérissé, A. and Wicander, R. 1996. Chapter 16. Paleozoic phytoplankton. In: Jansonius, J. and McGregor, D. C. (Eds.). Palynology: principles and applications. American Association of Stratigraphic Palynologists Foundation, 2: 495–529.
Ortega, G. and Albanesi, G. L. 2002. Bioestratigrafía de graptolitos y conodontes del Tremadociano tardío de la Cordillera Oriental argentina. In: Cabaleri N., Cingolani, C.A., Linares, E., López de Luchi, M.G., Ostera, H.A. y Panarello, H.O. (Eds.), Actas del XV Congreso Geológico Argentino CD–ROM, Artículo Nº 119, 6 pp.
Ottone, J.C., Toro, B.A. and Waisfeld, B.G. 1992. Lower ordovician palynomorphs from the Acoite formation, Northwestern Argentina. Palynology, 16: 93–116.
Ottone, E.G., Waisfeld, B.G. and Astini, R.A. 1995. Acritarcas del Ordovícico Temprano de la Quebrada de Chalala, Noroeste de Argentina. Ameghiniana, 32 (3): 237–242.
Playford, G., Ribecai, C. and Tongiorgi, M. 1995, Ordovician acritarch genera Peteinosphaeridium, Liliosphaeridium, and Cycloposphaeridium: morphology, taxonomy, biostratigraphy, and palaeogeographic significance. Bollettino della Società Paleontologica Italiana, 34 (1): 3– 54.
Raevskaya, E. G. 1999. Early Arenig acritarchs from the Leetse Formation (St. Petersburg region, northwest Russia) and their palaeogeographic significance. Bollettino della Società Paleontologica Italiana, 38 (2–3): 247–256.
Richardson, J. B. 1996. Chapter 18. Lower and middle Palaeozoic records of terrestrial palynomorphs. In: Jansonius, J. and McGregor, D. C. (Eds.). Palynology: principles and applications. American Association of Stratigraphic Palynologists Foundation, 2: 555–574.
Rubinstein, C.V. 1997. Tremadocian acritarchs from northwestern Argentina. Review of Palaeobotany and Palynology, 98: 41–45.
Rubinstein, C.V. 2001a. Correlaciones entre las cuencas ordovícicas y Silúricas de la Argentina basadas en el estudio de acritarcos, criptoesporas y mioesporas. In: Correlacao de Seqüencias Paleozóicas Sul–Americanas. Melo, J.H.G. and Terra, G.J.S. (Eds.). Ciencia–Técnica–Petróleo. Secao: Exploracao de Petróleo, 20: 19–24.
Rubinstein, C.V. 2001b. Bioestratigraphic and palaeogeographic implications of Lower Ordovician acritarchs from the Eastern Cordillera and Famatina, northwestern Argentina. Early Palaeozoic Palaeogeography and Palaeobiogeography of Western Europe and North Africa. Université des Sciences et Technologies de Lille (USTL), Lille. Abstracts: 57.
Rubinstein, C.V. 2003. Palinomorfos del Ordovícico y Silúrico en el Río Capillas, Sierras Subandinas, Argentina XII Simposio Argentino de Paleobotánica y Palinología. Resúmenes.
Rubinstein, C.V. and Astini, R.. 2000. Primer registro de palinomorfos arenigianos en las formaciones Suri y Molles, región del Famatina, provincia de La Rioja, Argentina. XI Simposio Argentino de Paleobotánica y Palinología. Resúmenes: 98.
Rubinstein, C.V. and Toro, B.A. 1999. Acritarch and graptolite biostratigraphy in the lower Arenig of the peri–Gondwana related Eastern Cordillera. Acta Universitatis Carolinae. (Geologica), 43 (1–2): 255–258.
Rubinstein, C.V. and Toro, B.A. 2001. Review of acritarch biostratigraphy in the Arenig of Eastern Cordillera, northwestern Argentina. New data and calibration with the graptolite zonation. In: Weiss R.H. (Eds.), Contributions to Geology and Palaeontology of Gondwana – In honour of Helmut Wopfner. Geological Institute, University of Cologne Germany: 421–439.
Rubinstein, C. and Toro, B., 2002 The messaoudensis–trifidum acritarch assemblage (late Tremadoc–early Arenig) from Parcha, Eastern Cordillera, northwestern Argentina. CIMP International Meeting and Workshops, Lille, France. Palaeozoic Palynology in the Third Millenium: new directions in acritarch, chitinozoan and miospore research. Abstract Volume: 48.
Rubinstein, C.V. and Vaccari, N.E. In press. Cryptospore assemblages from the Ordovician/Silurian boundary in the Puna, North–West Argentina. Palaeontology.
Rubinstein, C. V., Mángano, M.G. and Buatois, in press. Late Cambrian acritarchs from the Santa Rosita Formation: implications for the recognition of the Cambrian–Ordovician boundary in the Eastern Cordillera of northwest Argentina. Revista Brasileira de Paleontologia.
Rubinstein, C.V., Toro, B.A. and Waisfeld, B.G. 1999. Acritarch biostratigraphy of the upper Tremadoc – Arenig of the Eastern Cordillera, northwestern Argentina: relationships with graptolite and trilobite faunas. Bollettino della Società Paleontologica Italiana, 38 (2–3): 267–286.
Servais, T. and Molyneux, S. G. 1997, The messaoudensis– trifidum acritarch assemblage (Ordovician: late Tremadoc – early Arenig) from the subsurface of Rügen (Baltic Sea, NE Germany). Palaeontographia Italica, 84: 113– 161.
Tongiorgi, M. and Di Milia, A. 1999. Differentation and spread of the Baltic Acritarch Province (Arenig–Llanvirn). Bollettino della Società Paleontologica Italiana, 38 (2–3): 297–312.
Tongiorgi, M., Yin Lei–Ming and Di Milia, A., 1995, Arenigian acritarchs from the Daping section (Yangtze Gorges area, Hubei Province, Southern China) and their palaeogeographic significance. Review of Palaeobotany and Palynology, 86: 13– 48.
Toro, B. A. 1997. La fauna de graptolitos de la Formación Acoite, en el borde occidental de la Cordillera Oriental Argentina. Análisis bioestratigráfico: Ameghiniana, 34 (4): 393– 412.
Volkheimer, W., Melendi, D.L. and Aceñolaza, F.G. 1980. Una microflora ordovícica de la Formación Mojotoro, provincia de Salta. Revista de la Asociación Geológica Argentina, 35 (3): 401–416.
Received: February 15, 2003
Accepted: June 15, 2003