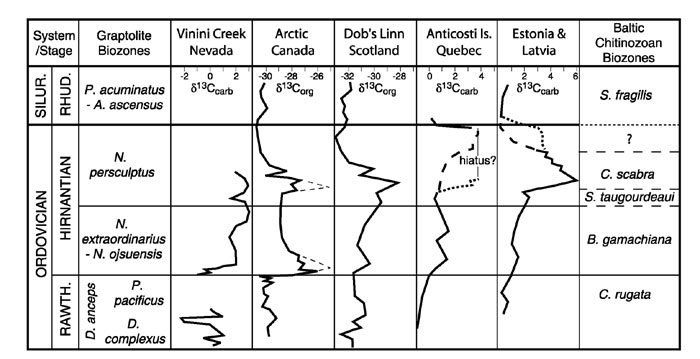
Correlation of graptolite biozones, chitinozoan biozones, and carbon isotope curves through the Hirnantian
Michael J. Melchin1, Chris Holmden2 and S. Henry Williams3
1 Department of Earth Sciences, St. Francis Xavier Unversity, Antigonish, Nova Scotia, B2G 2W5, Canada. E–mail: mmelchin@stfx.ca
2 Dept. of Geological Sciences, University of Saskatchewan, 114 Science Place, Saskatoon, SK S7N 5E2, Canada. E–mail: chris.holmden@usask.ca
3 Petro–Canada, 150 – 6th Avenue SW, Calgary AB, T3A 1Z2, Canada. E–mail: hwilliam@petro–canada.ca.
Key words: Graptolites. Chitinozoan. Biostratigraphy. Carbon isotope. Hirnantian.
New biostratigraphic and chemostratigraphic data are emerging from Dob’s Linn, Scotland, Anticosti Island, and Arctic Canada, that yield insights into the relative timing of the carbon isotope excursions and the graptolite biostratigraphy in those regions, as well as globally. These results cast doubt on the hypothesis that the timing and form of a composite curve based on the Baltic succession can be used a benchmark for global correlation through the Hirnantian (Brenchley et al., 2003).
At Dob’s Linn it has recently been discovered that Normalograptus persculptus occurs in what has previously been named the "extraordinarius Band" (Williams, 1983), 1.2 m below the base of the Birkhill shale. In addition, both N. extraordinarius and Normalograptus ojsuensis occur in the uppermost graptolitic strata of what had previously been regarded as the D. anceps Biozone ("anceps Band E" – Williams, 1982). Therefore, "anceps Band E", approximately 2.2 m below the base of the Birkhill Shale, should be regarded as the base of the N. extraordinarius–N. ojsuensis Biozone, which is the recently proposed base of the Hirnantian Substage (Rong et al., 1999). The base of the N. persculptus Biozone at this section must occur at or below the base of the "extraordinarius Band". The peak of the positive carbon isotope excursion at Dob’s Linn, which occurs at and immediately below the base of the Birkhill Shale (Underwood et al., 1997), is entirely within the N. persculptus Biozone, although C–isotope values begin to rise and show smaller peaks within the D. anceps and N. extraordinarius–N. ojsuensis Biozones (Figure 1).
On Anticosti Island graptolites previously assigned to Normalograptus angustus in the upper half the Ellis Bay Formation (Riva, 1988) have been reidentified as Normalograptus minor and Normalograptus parvulus, both species previously reported from strata not lower than the N. persculptus Biozone (Melchin, 2002). As at Dob’s Linn, this suggests that the most significant positive C–isotope excursion (Long, 1993) in the Anticosti succession is within the N. persculptus Biozone, although a weaker positive shift is also seen in the lower part of the Ellis Bay Formation, which, based on brachiopod data, appears to be lower Hirnantian (N. extraordinarius–N. ojsuensis Biozone) (Copper, 2001). These data suggest that, on Anticosti Island, the Belonechitina gamachiana Chitinozoan Biozone is lower Hirnantian (in the N. extraordinarius–N. ojsuensis Biozone ) and the Spinachitina taugourdeaui and Ancyrochitina ellisbayensis biozones are upper Hirnantian (in the N. persculptus Biozone). If these chitinozoan biozones are correlative between Anticosti Island and Baltica (Nõlvak, 1999), then the entire Porkuni Stage, the peak of the main positive C–isotope excursion and much the interval of rising C–isotope values observed in Estonia and Latvia (Brenchley et al., 2003) are also within the N. persculptus Biozone (Figure 1). In addition, if this correlation is correct, the peak occurrence of the Hirnantia Fauna in Baltica is also within the N. persculptus Biozone, as is demonstrably the case in northeastern Siberia (Koren’ et al., 1983). This correlation suggests that the lower Hirnantian interval, in the Baltic sections described by Brenchley et al. (2003), is highly condensed. Several of their described sections show discontinuity surfaces within or near the interval of the B. gamachiana Biozone.
Figure 1. Correlation of the carbon isotope curves with the graptolite and Baltic chitinozoan biozonation. Sources of carbon isotope data: Vinini Creek, Nevada – Finney et al. (1999); Arctic Canada – this study, composite of data from four sections (thin dashed line indicates high peak values seen at one section); Dob’s Linn, Scotland – Underwood et al. (1997); Anticosti Island – Long (1993), composite of data from two sections (dashed line indicate correlation supported by available biostratigraphic data, dotted line indicates correlation based on synchroneity of C–isotope profiles with a significant hiatus as proposed by Brenchley et al., 2003); Estonia and Latvia – Brenchley et al. (2003) (dashed line indicates correlation based on synchroneity of C–isotope profiles, dotted line is based on chitinozoan correlation assuming that base of S. fragilis Biozone coincides with base of Silurian).
In Arctic Canada, new C–isotope data from organic carbon successions show what is recognizable in several sections as two distinct positive excursions of comparable magnitude. The lower excursion begins at or near the base of the N. extraordinarius–N. ojsuensis Biozone (base of the Hirnantian). The upper excursion appears to reach its peak within the N. persculptus Biozone (Figure 1). A similar pattern and timing of two intervals of elevated C–isotope values occurs in South China (Wang et al., 1997). Whole–rock carbonate data from the same sections in Arctic Canada show more variable patterns of peaks, but a generally similar interval of overall positive values.
In Nevada (Finney et al., 1999) it appears that at least one phase of a major, positive C–isotope excursion begins within the N. extraordinarius Biozone and fluctuating but elevated values continue well into the N. persculptus Biozone (Figure 1).
At those sections that show apparently continuous deposition of graptolitic strata across the Ordovician–Silurian boundary (e.g., Dob’s Linn, Arctic Canada) the C–isotope values decline gradually through the upper N. persculptus Biozone and reach a low point at or just below the base of the Rhuddanian. On the other hand, at Anticosti Island, the positive C–isotope excursion appears, from biostratigraphic and lithostratigraphic evidence, to occur within the upper N. persculptus Biozone, and drops sharply at the boundary (Figure 1). It is not clear whether this difference between the Anticosti sections and graptolitic successions is the result of diachroneity in the faunal changes, a significant hiatus in the Anticosti succession, diachronous changes in the C–isotope geochemistry of the water masses, or a combination of these.
In Estonia and Latvia, the interval of declining C–isotope values occurs mainly within the S. fragilis Chitinozoan Biozone, which is generally regarded as lower Silurian (Brenchley et al., 2003). However, if the decline interval in Baltica is coincident with that seen in the graptolitic sections (Figure 1), then the base of the S. fragilis Biozone must be within the upper part of the Hirnantian.
Globally, it appears that the major interval of positive C–isotope excursions and eustatic fall extend from a level near the base of the lower Hirnantian N. extraordinarius–N. ojsuensis Biozone to a level within the N. persculptus Biozone (upper Hirnantian), and that C–isotope values decline through the upper Hirnantian strata. Within the d13Corg values, at least two distinct peaks can be resolved within the positive excursion interval, which may coincide with pulses of peak glaciation. In the regions around the Iapetus Ocean (e.g., Baltica, eastern Laurentia) the positive C–isotope excursions seem to be somewhat suppressed within the lower Hirnantian, and some may also shown differences in timing within the upper Hirnantian. This could be the result of differences in water chemistry within the Iapetus Ocean, which may have become relatively restricted in its connection with the world’s oceans during Hirnantian time as a result of the docking of Baltica with Avalonia and the progressive closure of the Iapetus Ocean (van Staal et al., 1998; Cocks and Torsvik, 2002). This restriction may also explain the survival of some endemic, relict benthic faunas through the Hirnantian in eastern Laurentia, Avalonia and Baltica (e.g., Owen and Robertson, 1995). Other differences between regions are also almost certainly the result of differing sedimentation rates, degrees of stratigraphic completeness, and diagenetic influences.
References
Brenchley, P.J., Carden, G.A., Hints, L. Kaljo, D., Marshall, J.D., Martma, T., Meidla, T., and Nõlvak, J. 2003. High–resolution stable isotope stratigraphy of Upper Ordovician sequences: constraints on timing of bioevents and environmental changes associated with mass extinction and glaciation. Geological Society of America Bulletin, 115: 89–104.
Cocks, L.R.M. and Torsvik, T.H. 2002. Earth geography from 500 to 400 million years ago: a faunal and paleomagnetic review. Journal of the Geological Society, London, 159: 631–644.
Copper, P. 2001. Reefs during the multiple crises towards the Ordovician–Silurian boundary: Anticosti Island, eastern Canada, and worldwide. Canadian Journal of Earth Sciences, 38: 153–171.
Finney, S.C., Berry, W.B.N., Cooper, J.D., Ripperdan, R.L., Sweet, W.C., Jacobson, S.R., Soufiane, A., Achab, A. and Noble, P.J. 1999. Late Ordovician mass extinction: A new perspective from stratigraphic sections in central Nevada. Geology, 27: 215–218.
Koren’, T.N., Oradovskaya, M.M., Pylma, L.J., Sobolevskaya, R.F., and Chugaeva. M.N. 1983. The Ordovician and Silurian boundary in the north–east of the USSR. Trudy Mezhvedomstvennogo Stratigraficheskogo Komiteta SSSR, 11. Nauka, Leningrad: 1–205 [In Russian].
Long, D.G.F. 1993: Oxygen and carbon isotopes and event stratigraphy near the Ordovician–Silurian boundary, Anticosti Island, Quebec. Palaeogeography, Palaeoclimatology, Palaeoecology, 104: 49–59.
Melchin, M.J. 2002. Restudy of some Ordovician–Silurian boundary graptolites from Anticosti Island: implications for the age and correlation of the Ellis Bay Formation. Canadian Paleontology Conference, Program and Abstracts Nº. 12, Ottawa, September, 2002: 31–33.
Nõlvak, J. 1999. Ordovician chitinozoan biozonation of Baltoscandia. Acta Universitatis Carolinae – Geologica, 43(1/2): 287–290.
Owen, A.W. and Robertson, D.B.R. 1995. Ecological changes during the end–Ordovician extinction. Modern Geology, 20: 21–39.
Riva, J. 1988: Graptolites at and below the Ordovician–Silurian boundary on Anticosti Island. In: Cocks, L.R.M. and Rickards, R.B. (Eds.): A global analysis of the Ordovician–Silurian boundary. Bulletin of the British Museum of Natural History (Geology), 43: 221–237.
Rong Jiayu, Chen Xu, Harper, D.A.T., and Mitchell, C.E. 1999. Proposal of a GSSP candidate section in the Yangtze Platform region, S. China, for a new Hirnantian boundary stratotype. Acta Universitatis Carolinae – Geologica, 43(1/2): 77–80.
Underwood, C.J., Crowley, S.F., Marshall, J.D., and Brenchley, P.J. 1997. High–resolution carbon isotope stratigraphy of the basal Silurian Stratotype (Dob’s Linn, Scotland) and its global correlation. Journal of the Geological Society, London, 154: 709–718.
van Staal, Dewey, J.F., MacNiocaill, C., and McKerrow, W.S. 1998. The Cambrian–Silurian tectonic evolution of the northern Appalachians and British Caledonides: history of a complex, west and southwest Pacific–type segment of Iapetus. In: Blundell, D.J. and Scott, A.C. (Eds.), Lyell: the Past is the Key to the Present, Geological Society, London, Special Publications, 143: 199–242.
Wang Kun, Chatterton, B.D.E. and Wang Y. 1997: An organic carbon isotope record of Late Ordovician to Early Silurian marine sedimentary rocks, Yangtze Sea, South China: implications for CO2 changes during the Hirnantian glaciation. Palaeogeography, Palaeoclimatology, Palaeoecology 132, 147–158.
Williams, S.H. 1982. The Late Ordovician graptolite fauna of the Anceps Bands at Dob’s Linn, southern Scotland. Geologica et Palaeontologica, 16, 29–56.
Williams, S.H. 1983. The Ordovician–Silurian boundary graptolite fauna of Dob’s Linn, southern Scotland. Palaeontology, 26, 605–639.
Received: February 15, 2003
Accepted: June 15, 2003