Acritarchs from peri–Gondwana at the Lower and Middle Ordovician Stage boundaries
Jun Li1, Stewart G. Molyneux2, Claudia V. Rubinstein3 and Thomas Servais4
1 Nanjing Institute of Geology and Palaeontology, Academia Sinica, Chi–Ming–Ssu, Nanjing 210008, China. E–mail: junli@nigpas.ac.cn
2 British Geological Survey, Keyworth – Nottingham, NG12 5GG, UK E–mail: sgm@bgs.ac.uk
3 IANIGLA – CRICYT C. C. 131 5500 – Mendoza, Argentina. E–mail: crubinstein@arlinkbbt.com.ar
4 UMR 8014 du CNRS, USTL, Cité Scientifique, 59655 Villeneuve d’Ascq Cedex, France. E–mail: thomas.servais@univ–lille1.fr
Key words: Acritarchs. Palynomorphs. Stage boundaries. Ordovician. Gondwana.
Introduction
Although acritarchs have been studied mostly for stratigraphic purposes (often in sediments devoid of other fossils), their biostratigraphic potential has not yet been fully exploited in the Ordovician. While another group of palynomorphs, the chitinozoans, have become increasingly important in international correlations, the acritarchs continue to play a minor role in Ordovician biostratigraphy.
Today, about 1000 papers deal with Ordovician acritarchs, most of which are published by scientists working in Europe. The geographical areas studied in detail are in Europe and North Africa (Servais et al., in press). Investigations of Laurentian acritarchs are generally limited to the upper Middle Ordovician and the Upper Ordovician. Apart from two publications with detailed descriptions, studies from Australia are almost non–existant. In recent years, an increasing number of investigations have been carried out in China (Li et al., 2002) and South America (e.g., Rubinstein and Toro, 2001). However, at a global scale, many areas remain almost unstudied.
Palaeobiogeographically, two distinct geographic acritarch assemblages can be recognized in the Lower–Middle Ordovician. A first assemblage (also named "province"), in the upper Tremadocian in low latitude areas, was attributed to warm–water environments, and a second assemblage, successively named "Mediterranean" or "peri–Gondwanan province", can easily be recognized around the southern border of Gondwana, from Argentina through north Africa and peri–Gondwana to Iran, Pakistan and southern China (Vavrdová, 1974; Li and Servais, 2002).
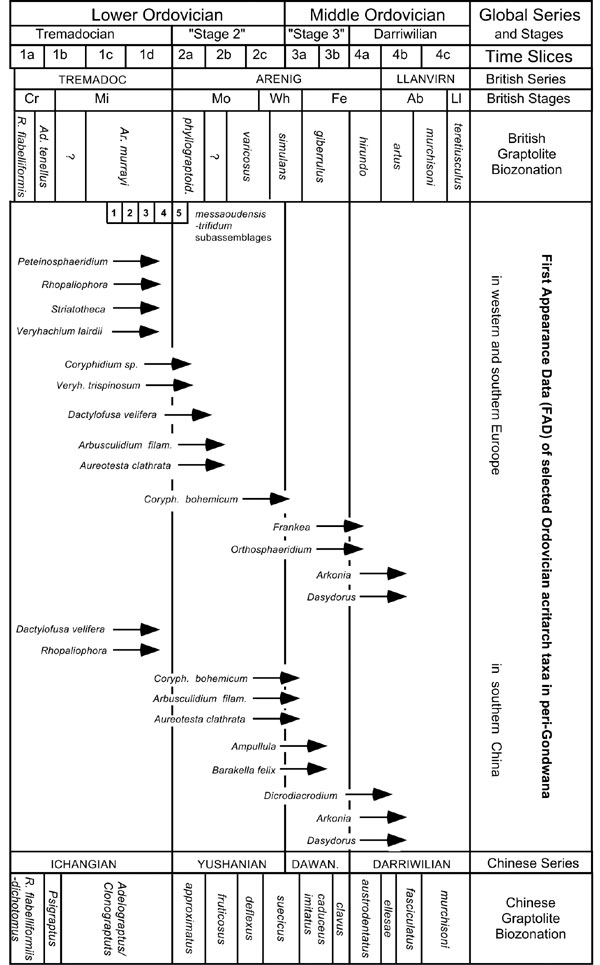
Over 250 genera and over 1500 species of Ordovician acritarchs have been described so far (Servais et al., in press). Among the more than 500 acritarch species described from peri–Gondwana, some may be used as stratigraphical index taxa and may be helpful in the recognition and correlation of the Global Stage boundaries in the Lower and Middle Ordovician. The first appearance datum (FAD) of some 17 taxa, the taxonomy of which has been thoroughly reviewed in recent years, are plotted in this study against the British and Chinese graptolite zonations (Figure 1), following similar treatment of the FADs of 8 taxa by Brocke et al. (1995). The last appearance datum (LAD) of these taxa is more problematical, as palynomorphs of the size of the acritarchs may be reworked into younger sediments. LADs may be suggested with confidence in some instances, but only after a consistent pattern of occurrence has been determined from large data sets.
The Tremadocian–"Stage 2" boundary
Although recognition of the Cambrian–Ordovician boundary using acritarchs remains problematical (e.g., Vecoli, 1999; Parsons and Anderson, 2000), recognition of the Tremadocian–"Stage 2" boundary based on acritarchs is now possible, following investigations in continuous sections with graptolite and/or chitinozoan control from the British Isles, Germany, the Czech Republic and Spain (for a review of the literature, see Servais and Mette, 2002). Following the original description of the messaoudensis–trifidum acritarch assemblage from the English Lake District (Molyneux and Rushton, 1988), Cooper et al. (1995) subdivided the easily recognizable acritarch assemblage into five sub–assemblages, informally designated 1 to 5 upwards. Three typical Ordovician acritarch genera, Peteinosphaeridium, Rhopaliophora and Striatotheca, appear for the first time in sub–assemblage 1, corresponding to the lower part of the A. murrayi graptolite Biozone, together with the rectangular specimens of Veryhachium (V. lairdii group). The first specimens of triangular Veryhachium (V. trispinosum group) and of the genus Coryphidium appear higher in the successions, but below the Tremadocian–"Stage 2" boundary (Cooper et al., 1995; Molyneux, unpublished data). Dactylofusa velifera also appears below the first appearance of T. phyllograptoides, and thus its FAD is still in the Tremadocian (Servais and Mette, 2000). The species Arbusculidium filamentosum and Aureotesta clathrata, however, first appear above the boundary in the sub–assemblage 5 of Cooper et al. (1995). The FADs of these two taxa, reviewed by Fatka and Brocke (1999) and Brocke et al. (1998), respectively, are thus indicators for the lowermost part of the "Global Stage 2", or, in terms of British stratigraphy, for the lowermost Arenig.
The South Chinese acritarch succession provides only poorly diverse assemblages for this time interval (Li in Servais et al., in press). Nevertheless, the presence of the species Dactylofusa velifera and the genus Rhopaliophora in the upper Tremadocian has been confirmed in the Yangtze Platform (Brocke, 1997). Acritarch investigations from the Tremadocian–"Stage 2" interval in the Eastern Cordillera of Argentina indicate that some typical forms of the messaoudensis–trifidum acritarch assemblage, including the genus Coryphidium, are present in strata related to the A. murrayi graptolite Biozone, confirming the first appearance of this genus in the upper Tremadocian (Rubinstein and Toro, 2002).
The Lower–Middle Ordovician boundary
Li et al. (2002) have recently documented the potential of acritarchs from the South Chinese Yangtze Platform in the recognition of the Lower–Middle Ordovician boundary. Well–preserved and highly diverse acritarch assemblages have been recovered from numerous sections in the Yangtze Platform and the Jianshan–Changshan–Yushan (JCY) area, crossing the stratigraphic interval in which the Lower–Middle Ordovician boundary should be located. The highest diversities are observed in the D. deflexus, A. suecicus and I. caduceus imitatus graptolite Biozones, where a succession of FADs of easily distinguishable acritarch taxa can be observed. Coryphidium bohemicum, Arbusculidum filamentosum and Aureotesta clathrata are all present from the D. deflexus graptolite Biozone, whereas the genus Ampullula and the species Barakella felix first appear only in the A. suecicus Biozone, thus indicating the boundary interval (Li et al., 2002). Acritarch assemblages from the B. deflexus and D. bifidus (approximately equivalent to the A. suecicus) graptolite Biozones are also very rich and well documented in northwestern Argentina, where Coryphidium bohemicum, Arbusculidum filamentosum and Aureotesta clathrata also occur. The occurrence of Barakella in the D. bifidus graptolite Biozone suggests, as in South China, the proximity of the Lower–Middle Ordovician boundary (Rubinstein and Toro, 2001).
Figure 1
Most of these occurrences are of great importance for an understanding of acritarch biostratigraphy in peri–Gondwana, since most taxa are widespread in all areas on the southern border of Gondwana.
The "Stage 3"–Darriwilian boundary
The recognition of this boundary is also possible with the help of the FADs of selected acritarch taxa. The widely distributed Ordovician acritarch genera Frankea and Orthosphaeridium appear for the first time in the I. gibberulus graptolite Biozone, whereas Arkonia and Dasydorus only appear in the D. hirundo graptolite Biozone in the British Isles (Servais, 1993; Cooper et al., 1995; Servais, 1997). The latter two genera also appear at precisely the same time in China, in the U. austrodentatus graptolite Biozone, i.e., at the base of the Darriwilian (Li et al., 2002). These two genera are thus very good indicators for the base of the Darriwilian, the fourth global stage of the Ordovician.
Additionally, the genus Dicrodiacrodium, which occurs widely along the southern border of Gondwana, has its FAD in the southern Chinese sections just below the "Stage 3"–Darriwilian boundary in the E. clavus graptolite Biozone.
Studies on Middle and Upper Ordovician acritarchs from the Subandean Ranges, northwestern Argentina, have only recently begun. Here, the genus Arkonia has been recorded in the upper Llanvirn (Rubinstein, 2003).
Conclusions
A succession of first appearance data (FAD) for 17 easily recognizable acritarch taxa, the taxonomy of which has been reviewed recently, are identified in assemblages ranging from the upper Tremadocian to the lowermost Darriwilian of peri–Gondwana. The taxa belong to an acritarch "province" that extends around the southern border of Gondwana, from Argentina in the east to southern China in the west. FADs of individual taxa throughout this geographic range suggest that acritarchs can contribute to the correlation of global Ordovician series and stage boundaries, at least on the margin of peri–Gondwana.
References
Brocke, B. 1997. First results of Tremadoc to lower Arenig acritarchs from the Yangtze Platform, southwest China. In : Fatka O. and Servais T. (Eds.), Acritarcha in Praha, Acta Universitatis Carolinae, Geologica, 40: 337–355.
Brocke, R., Fatka, O., Molyneux, S.G. and Servais T. 1995. First appearance of selected early Ordovician acritarch taxa from peri–Gondwana. In: Cooper C., Droser, M.L. and Finney S.C. (Eds.), Ordovician Odyssey. The Pacific Section for Sedimentary Society (SEPM), Fullerton, California, USA, 77: 473–476.
Brocke, R., Fatka, O. and Servais T. 1998. A review of the Ordovician acritarchs Aureotesta and Marrocanium. Annales de la Société Géologique de Belgique, 120(1): 1–21.
Cooper, A.H., Rushton, A.W.A., Molyneux, S.G., Hughes, R.A., Moore, R.M. and Webb, B.C. 1995. The stratigraphy, correlation, provenance and palaeogeography of the Skiddaw Group (Ordovician) in the English Lake District. Geological Magazine, 132: 185–211.
Fatka, O. and Brocke R. 1999. Morphologic variability in two populations of Arbusculidium filamentosum (Vavrdová 1965) Vavrdová 1972. Palynology, 23: 153–180.
Li, J. and Servais, T. 2002. Ordovician acritarchs of China and their utility for global palaeobiogeography. Bulletin de la Société Géologique de France, 173(5): 399–406.
Li, J., Brocke, R. and Servais, T. 2002. The acritarchs of the South Chinese Azygograptus suecicus graptolite Biozone and their bearing on the definition of the Lower/Middle Ordovician boundary. C.R. Palevol, 1: 75–81.
Molyneux, S.G. and Rushton, A.W.A. 1988. The age of the Watch Hill Grits (Ordovician), English Lake District: structural and palaeogeographical implications. Transactions of the Royal Society of Edinburgh, Earth Sciences, 79: 43–69.
Parsons, M.G. and Anderson M.M., 2000. Acritarch microfloral succession from the Late Cambrian and Ordovician (early Tremadoc) of Random Island, eastern Newfoundland, and its comparison to coeval microfloras, particularly those of the East European Platform. American Association of Stratigraphic Palynologists Foundation, Contribution Series, 38: 1–123.
Rubinstein, C.V., 2003. Palinomorfos del Ordovícico y Silúrico en el Río Capillas, Sierras Subandinas, Argentina. XII Simposio Argentino de Paleobotánica y Palinología. Resúmenes.
Rubinstein, C.V. and Toro, B.A. 2001. Review of acritarch biostratigraphy in the Arenig of Eastern Cordillera, northwestern Argentina. New data and calibration with the graptolite zonation. In: Weiss R.H. (ed.), Contributions to Geology and Palaeontology of Gondwana – In honour of Helmut Wopfner. Geological Institute, University of Cologne, Germany: 421–439.
Rubinstein, C.V. and Toro, B.A. 2002. The messaoudensis–trifidum acritarch assemblage (late Tremadoc–early Arenig) from Parcha, Eastern Cordillera, northwestern Argentina. In: Raevskaya, E. and Servais, T. (Eds.), CIMP International Meeting and Workshops, Lille, France. Palaeozoic Palynology in the Third Millenium: new directions in acritarch, chitinozoan and miospore research. Abstract Volume, p. 48.
Servais, T. 1993. The Ordovician acritarch Frankea. Special Papers in Palaeontology, 48: 79–95.
Servais, T. 1997. The Ordovician Arkonia–Striatotheca acritarch plexus. Review of Palaeobotany and Palynology, 98: 47–79.
Servais, T. and Mette, W. 2002. The messaoudensis–trifidum acritarch assemblage (Ordovician: late Tremadoc–early Arenig) of the Barriga Shale Formation, Sierra Morena (SW–Spain), Review of Palaeobotany and Palynology, 113: 145–163.
Servais, T., Li, J., Stricanne, L., Vecoli, M., and Wicander, R. (in press). Chapter 32. Acritarchs. In: Webby, B., Droser, M., Paris, F. and Percival, I. (Eds.), The Great Ordovician Biodiversification Event. Columbia University Press.
Vavrdová, M. 1974. Geographical differentiation of Ordovician acritarch assemblages in Europe. Review of Palaeobotany and Palynology, 18: 171–176.
Vecoli, M. 1999. Cambro–Ordovician palynostratigraphy (acritarchs and prasinophytes) of the Hassi–R’Mel area and northern Rhadames Basin, North Africa. Palaeontographia Italica, 86: 1–112.
Received: February 15, 2003
Accepted: June 15, 2003