The Diplacanthograptus caudatus Zone: a proposal for a new, globally recognizable, Upper Ordovician graptolite zone
Daniel Goldman1
1 Dept. of Geology, University of Dayton, 300 College Park, Dayton, OH 45469 . E–mail: dan.goldman@notes.udayton.edu
Key words: Graptolites. Biozone. Biostratigraphy. Correlation. Ordovician.
Introduction
The Ordovician Sub–commission of the International Commission on Stratigraphy has recently redefined the base of the Upper Ordovician Series as the base of the Nemagraptus gracilis graptolite biozone. This designation recognized the tremendous utility of a globally identifiable biozone based on the first appearance of a cosmopolitan taxon that occurs within a consistent succession of other first appearance datums (FADs) (e.g., Finney and Bergström, 1986; Bergström et al., 2000). In the upper part of the range of N. gracilis, Climacograptus bicornis, members of the Orthograptus calcaratus species group, and a profusion of large dicellograptinids and dicranograptinids first appear (Finney and Bergström, 1986). This fauna is also widely known and can be precisely correlated across the world.
Unfortunately, the correlation of post–Climacograptus bicornis Zone rocks is one of the long–standing problems in Ordovician graptolite biostratigraphy. Faunal provincialism has led to the construction of numerous separate zonations, 4 in North America alone. Establishing detailed correlations between these various zonations has proved difficult and often contentious (e.g., Riva, 1969, 1974; Berry 1970, 1977; Bergström, 1978; Finney, 1986). In this paper I will briefly review the post–Climacograptus bicornis Zone graptolite successions from several well known localities and suggest a more useful standardized zonation for this interval based on the Climacograptus – Diplacanthograptus lineages.
Review of post Climacograptus bicornis Zone graptolite successions
Australasia.
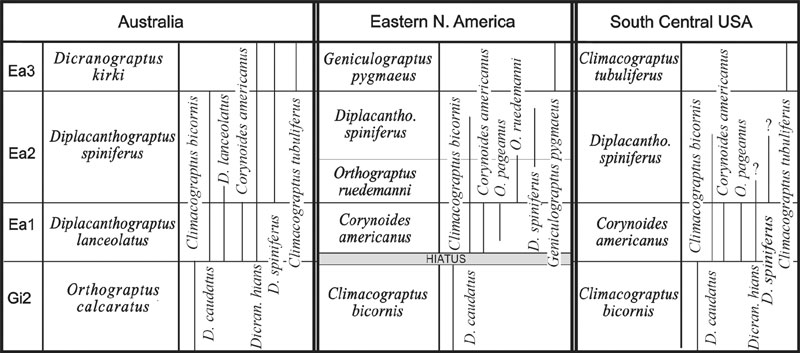
The Australasian graptolite sequence is one of the most complete and finely divided sequences in the world. It is considered to be the standard for the Pacific Faunal Province (VandenBerg and Cooper, 1992). The first post–C. bicornis biozone is the Diplacanthograptus lanceolatus Zone (Ea1, see Figure 1a). This zone is characterized by the appearance of D. lanceolatus, D. caudatus, Corynoides americanus and Dicranograptus hians (VandenBerg and Cooper, 1992). The D. lanceolatus Zone is succeeded by the Diplacanthograptus spiniferus Zone (Ea2), and the Dicranograptus kirki Zone (Ea3). Correlation of Ea1 and Ea2 strata with other graptolite successions is complicated by the fact that the nominal species, D. lanceolatus, is not widely known outside of Australia and by the globally diachronous appearance of D. spiniferus (Figure 1a, b). The Dicranograptus kirki Zone can be recognized in most regions (excluding eastern North America) by the presence of D. kirki and Climacograptus tubuliferus.
Figure 1a. Correlation of post–Nemagraptus gracilis Upper Ordovician graptolite biozones with a comparison of the stratigraphic ranges of some key index species. The chart is continued in Figure 1b on the facing page. Data on graptolite ranges from Finney (1986), Goldman et al. (1999), Riva, (1969, 1974), and VandenBerg and Cooper (1992).
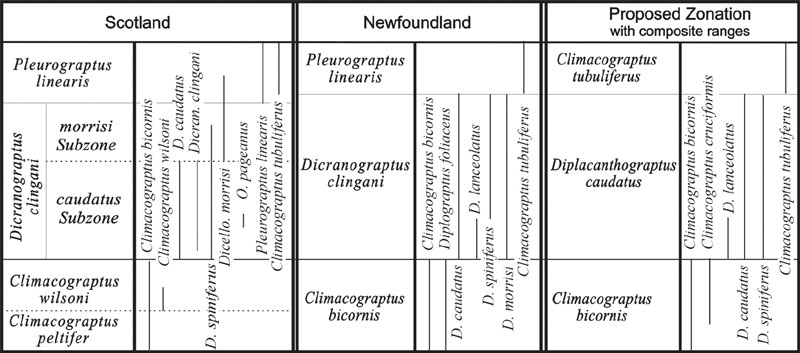
Figure 1b. Continuation of correlation chart in Figure 1a. Note that the last column depicts a proposed new biozone based on the range of Diplacanthograptus caudatus. Data on graptolite ranges from VandenBerg and Cooper (1992), Williams (1995), and Zalasiewicz et al. (1995).
Eastern North America. The classical New York State–Quebec graptolite sequence described by Ruedemann (1908, 1912, 1925) and Riva (1969, 1974) contains one of the best known and most provincial faunas in post–C. bicornis Zone rocks. There are a large number of endemic species and many FADs are late with respect to their position elsewhere in the world. Notable late arrivals into the Appalachian Basin are D. spiniferus, O. spinigerus, and O. pageanus (Goldman et al., 1999). One of the most striking characteristics of the C. bicornis – C. americanus zonal boundary in North America is a great reduction in species diversity. Nearly all the dicellograptinids, dicranograptinids, and members of the Orthograptus calcaratus species group disappear at the base of the C. americanus Zone. Many of these taxa occur abundantly in this interval elsewhere in the world and inter–continental correlations in Upper Ordovician strata remain problematic. C. americanus Zone strata can be correlated with the Australasian sequence by the presence of C. americanus, O. pageanus, and D. caudatus. D. lanceolatus does not occur in the New York – Quebec region. Correlations with other regions (e.g., Scotland and Newfoundland) can be made based on the presence of D. caudatus. Above the C. americanus Zone, faunas in eastern North America become increasingly endemic and difficult to correlate (Riva, 1974; Goldman et al., 1995).
South Central United States. The graptolite succession of the Arbuckle and Ouachita mountains of Central Oklahoma has recently been revised by Finney (1986). The fauna contains a mixture of typical Pacific Province taxa and Laurentian endemics. Additionally, the hiatus that exists between the C. bicornis and C. americanus zones in most Eastern North American sections (Figure 1a) is not present in Oklahoma. Thus, the Oklahoma succession provides a useful intermediary for correlating pacific Province localities with the New–York Quebec succession. The first appearance of D. caudatus, Dicranograptus hians, C. americanus, and O. pageanus mark the base of the C. americanus Zone and suggest precise correlations with localities outside of North America.
A large unconformity spans this interval (Ea1 – Ea3) in the Marathon Region of West Texas (Riva, 1974; Bergström, 1978; Goldman et al., 1995).
Scotland. The graptolite sequence in southern Scotland is historically one of the most important and well documented successions in the world (Lapworth 1876; Elles and Wood, 1901 – 1918). More recently, it has been thoroughly revised by Williams (1982) and Zalasiewicz et al., (1995). Little can be added here except to note that Zalasiewicz et al., (1995) recognized the utility of sub–dividing the Dicranograptus clingani Zone into a caudatus subzone and a morrisi subzone for correlation purposes. They also noted that the overlying Pleurograptus linearis Zone is best recognized not by the nominal species, which is restricted to very few horizons and it is difficult to identify in fragmentary material, but by the presence of Climacograptus styloideus and Climacograptus tubuliferus.
Newfoundland. Ordovician graptolites from the Lawrence Harbour Formation in north–central Newfoundland have been described by Erdtmann (1976) and Williams (1995). The graptolite succession is most similar to that of Australia, although Williams (1995) adopted the British zonal scheme. It is important to note that while Williams uses the Scottish zonation, he recognizes the D. clingani Zone based primarily on the presence of D. caudatus, D. lanceolatus, D. spiniferus; and the P. linearis Zone based on the occurrence of Climacograptus tubuliferus.
Proposal for a new globally recognizable biozone
It is clear from the brief outline of post–C. bicornis Zone biostratigraphy presented above that increased faunal provincialism has resulted in the construction of numerous different zonations. Establishing precise correlations between these regional zonations has proved to be difficult and contentious. Yet a close inspection of the faunas present in each region reveals a single taxon, Diplacanthograptus caudatus, which is an abundant element of each fauna, has a consistent FAD within a succession of first appearances by other taxa, and is easily recognizable. Additionally, new isolated three–dimensionally preserved material of D. caudatus (Goldman and Wright, this volume) has clarified the proximal end morphology and phylogenetic relationships of this taxon. Diplacanthograptus caudatus is part of a well studied lineage comprising C. bicornis (Hall) – D. lanceolatus (VandenBerg) – D. caudatus (Lapworth) – D. spiniferus (Ruedemann) and C. tubuliferus Lapworth (VandenBerg, 1990).
In one anomalous report, Nilsson (1977) identified specimens of C. caudatus in much older strata (H. teretiusculus Zone) in the Koängen Core from Scania, southern Sweden. Those specimens were misidentified and belong to an undescribed species of Proclimacograptus Maletz.
Thus, one possible solution to the biostratigraphic problem discussed above is a new biozone based on the range of Diplacanthograptus caudatus, which occurs abundantly in nearly all the described graptolite faunas from the Upper Ordovician. Other characteristic but not ubiquitous components of this zone are D. lanceolatus, Corynoides americanus, Orthograptus pageanus, O. quadrimucronatus, Dicranograptus hians, and Neurograptus margaritatus. A D. caudatus Zone is more useful than the D. lanceolatus Zone because the latter taxon is not common outside of Australia. The D. caudatus Zone would succeed the Climacograptus bicornis Zone and, in turn, be overlain by a C. tubuliferus Zone. The commonly utilized D. spiniferus Zone should be restricted to local usage because of the highly diachronus FAD of its nominal species.
References
Bergström, S. M. 1978. Middle and Upper Ordovician conodont and graptolite biostratigraphy of the Marathon, Texas graptolite zone reference standard. Palaeontology 21: 723–758.
Bergström, S. M., et al., 2000. A proposed global boundary stratotype for the base of the Upper Series of the Ordovician System: the Fågelsång section, Scania, southern Sweden. Episodes 23: 102–109.
Berry, W.B.N. 1970. Review of late Middle Ordovician graptolites in eastern New York and Pennsylvania. American Journal of Science, 269: 304–313.
Berry, W.B.N. 1977. Ecology and age of graptolites from graywackes in eastern New York. Journal of Paleontology 51: 1102–1107.
Elles G.L. and Wood E.M.R. 1901–1918. A monograph of British graptolites. Palaeontographical Society Monographs, clxxi + 539.
Erdtmann, B.–D. 1976. Die Graptolithenfauna der Exploits–Gruppe von Zentral–Neufundland. Mitteilungen aus dem Geologisch–Paläontologischen Institut der Universität Hamburg, 45: 65–140.
Finney, S.C. 1986. Graptolite biofacies and correlation of eustatic, subsidence, and tectonic events in the Middle to Upper Ordovician of North America. Palaios, 1: 435–461.
Finney, S.C. and Bergström, S.M. 1986. Biostratigraphy of the Ordovician Nemagraptus gracilis Zone, In C.P. Hughes and R.B. Rickards Eds., Paleoecology and biostratigraphy of graptolites. Geological Society Special Publication, 20: 47–59.
Goldman, D., Bergström, S.M., and Mitchell, C.E. 1995. Revision of the Zone 13 graptolite biostratigraphy in the Marathon, Texas, standard succession and its bearing on Upper Ordovician graptolite biogeography. Lethaia, 28: 115–128.
Goldman, D. Mitchell, C.E., Joy, M.P. 1999. The stratigraphic distribution of graptolites in the classic Middle Ordovician Utica Shale of New York State: an evolutionary succession or a response to relative sea–level change? Paleobiology, 25: 273–294.
Lapworth, C. 1876. The Silurian System in the south of Scotland. In Armstrong, J., Young, J., and Robertson, D. Catalogue of Western Scottish Fossils: 1–28. Blackie and Son, Glasgow.
Nilsson, R. 1977. A boring through Middle and Upper Ordovician strata at Koängen in western Scania, southern Sweden. Sveriges Geologiska Undersökning, C733: 1–58.
Riva, J. 1969. Middle and Upper Ordovician graptolite faunas of St. Lawrence Lowlands of Quebec, and of Anticosti Island. In M. Kay, ed. North Atlantic – geology and continental drift. A symposium. American Association of Petroleum Geologists Memoir, 112: 579–595, Tulsa, Oklahoma.
Riva, J. 1974. A revision of some Ordovician graptolites of eastern North America. Palaeontology, 17:1–40.
Ruedemann, R. 1908. Graptolites of New York. Part II. Graptolites of the higher beds. New York State Museum Memoir 11, 547.
Ruedemann, R. 1912. The Lower Siluric shales of the Mohawk Valley. New York State Museum Bulletin, 162, 151.
Ruedemann, R. 1925. The Utica and Lorraine formations of New York, Pt. 1, Stratigraphy: New York State Museum Bulletin, 258, 174 .
VandenBerg, A.H.M. 1990. The ancestry of Climacograptus spiniferus Ruedemann. Alcheringa, 14: 39–51.
VandenBerg, A.H.M., and Cooper, R.A. 1992. The Ordovician graptolite sequence of Australasia. Alcheringa, 16: 33–85.
Williams, S.H. 1982. Upper Ordovician graptolites from the top Lower Hartfell Shale Formation (D. clingani and P. linearis zones) near Moffat, southern Scotland. Transactions of the Royal Society of Edinborough, Earth Sciences, 72: 229–255.
Williams, S.H. 1995. Middle Ordovician Graptolites from the Lawrence Harbour Formation, Central Newfoundland, Canada. Palaeontographica Abt. A, 235: 21–77.
Zalasiewicz, J., Rushton, A.W.A., and Owen, A.W. 1995 Late Caradoc graptolitic faunal gradients across the Iapetus Ocean. Geological Magazine, 132: 611–617.
Received: February 15, 2003
Accepted: June 15, 2003