The Red River problem revisited: stratigraphic relationships in the Upper Ordovician of central and western United States
Stig M. Bergström1
1 Department of Geological Sciences, The Ohio State University, Columbus, OH 43210, USA. E–mail: stig@geology.ohio-state.edu
Key words: Red River. Stratigraphy. Environments. Upper Ordovician. USA.
Introduction
One of the most controversial, and much discussed, problems in the North American Ordovician geology is the age of a prominent carbonate unit that forms the topmost part of the system in many regions. It can be traced from the Canadian Arctic to Texas, and from the eastern Midcontinent to the Great Basin and contains a distinctive shelly fossil assemblage long known as the Red River (Arctic) fauna. From the 1920s to the present, the rocks having this fauna have been regarded as late Middle Ordovician (late Mohawkian), or late Late Ordovician (Richmondian), or representing a large stratigraphic interval in the Late Ordovician (Cincinnatian). The present paper evaluates biostratigraphic data pertaining to this carbonate unit in an effort to establish its age locally and regionally.
Biostratigraphic framework
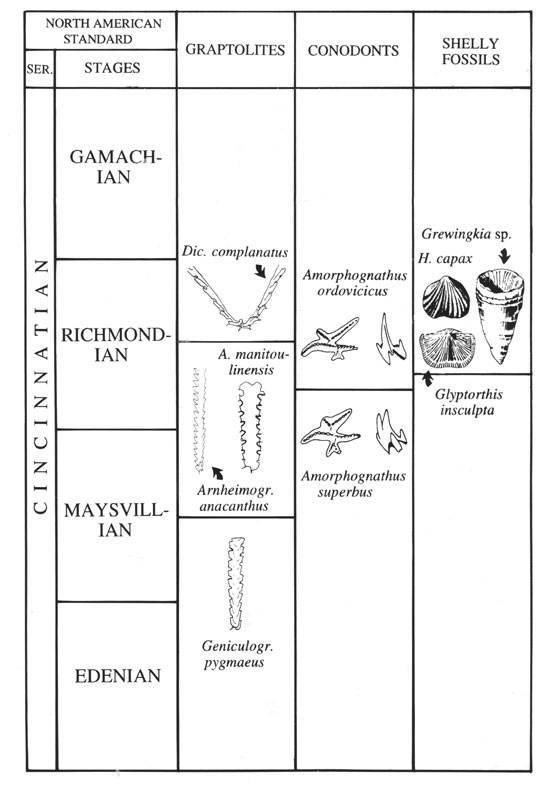
The Cincinnatian Series has traditionally been subdivided into the Edenian, Maysvillian, Richmondian, and Gamachian Stages. A succession of conodonts largely based on sections in the Cincinnati region was introduced by Sweet (1984). An alternative zone scheme based mainly on Baltoscandic sections was proposed by Bergström (1971). Of particular interest for the present study is the base of the A. ordovicicus Zone, which in the Cincinnati region is in the lower, but not lowermost, Richmondian (MacKenzie and Bergström, 1994; Figure 1). All Pre–Richmondian Amorphognathus occurrences in the type Cincinnatian represent A. superbus (Goldman and Bergström, 1997). The type Cincinnatian ranges from the upper C. spiniferus Zone through the A. manitoulinensis Graptolite Zone (Bergström and Mitchell, 1986) but much of the Richmondian lacks graptolites, and the Gamachian is missing. The youngest Ordovician graptolite zones are best known from central Nevada (Finney et al., 1999). Achab (l989) and Soufiane and Achab (2000) proposed 12 chitinozoan zones through the Cincinnatian and correlated these with the graptolite succession. The base of the A. ordovicicus Zone corresponds to an unzoned chitinozoan interval between the Achab’s (1989) C. hyalophrys and C. senta Zones. In the type Cincinnatian, the Hiscobeccus capax–Grewingkia robusta fauna (in the following abbreviated Hiscobeccus–Grewingkia fauna), which is a significant component of the Red River or Arctic fauna, is especially important biostratigraphically. This very widespred fauna, which includes a distinctive assemblage of brachiopods, gastropods, corals, and cephalopods, appears in the middle Richmondian in the Cincinnati region well above the base of the A. ordovicicus Zone, in strata corresponding to a level near the A. manitoulinensis–D. complanatus Zone boundary (Figure 1).
Figure 1. Stratigraphic position of some Cincinnatian key taxa discussed in the text.
Regional stratigraphic review
In the well exposed and stratigraphically apparently complete Cincinnatian succession in the Arbuckle Mountains of Oklahoma, the base of the A. ordovicicus Zone is in the upper Viola Springs Formations somewhat below the base of the Welling Formation in an interval referable to the A. manitoulinensis Zone (Goldman and Bergström, 1997). The Hiscobeccus–Grewingkia fauna occurs in the Welling Formation, which is overlain by the graptolitiferous Sylvan Shale which has the base of the D. complanatus Zone in its lowermost part (Goldman and Bergström, 1997). Hence, the relations between conodonts, graptolites, and shelly faunas are the same as in the type Cincinnatian, and the upper part of the Viola Springs, the Welling, and the Sylvan Shale clearly represent the Richmondian Stage (Figure 2).
In the classical Marathon sequence, West Texas, the basal part of the Maravillas Formation contains conodonts of the A. ordovicicus Zone, and graptolites of the D. gravis Zone. This zone is overlain by the D. ornatus Zone (Goldman et al., 1995) that has yielded some shelly fossils suggesting the Hiscobeccus–Grewingkia fauna and the Maravillas is clearly of Richmondian age. In the Montoya Group of New Mexico and adjacent part of West Texas, the upper part of the Second Value Formation, and the overlying Aleman Formation, have yielded a Hiscobeccus–Grewingkia fauna (Howe, 1959) as well as a diverse A. ordovicicus Zone conodont fauna (Sweet, 1979). Athough no graptolites are known from the Montoya, this unit, which is generally correlated with the Maravillas, is also likely to be of Richmondian age.
The base of the A. ordovicicus Zone is in the Cape Formation of southeastern Missouri and in the uppermost Dubuque Formation of Iowa (Goldman and Bergström, 1997). The Dubuque is overlain by the Maquoketa Shale, the lower part of which contains graptolites of the A. manitoulinensis Zone. Higher parts of the Maquoketa Shale (Ft. Atkinson Formation) in Illinois has yielded the graptolite zonal index D. complanatus, and the Maquoketa also contains a good representation of the Hiscobeccus–Grewingkia fauna. As shown by Goldman and Bergström (1997), the Maquoketa of Iowa can be closely correlated with the type Richmondian succession in the Cincinnati region.
In the Western Interior, the diverse conodont fauna of the Bighorn Group of Wyoming (Sweet, 1979) is similar to that of the Fremont and Priest Canyon Formations of Colorado. The upper Bighorn contains the Hiscobeccus–Grewingkia fauna (Macomber, 1970) but the basal part of the Wyoming and Colorado units has not yet yielded A. ordovicicus although the rest of the conodont fauna is quite similar to that present in, for instance, the lower part of the Montoya Group. Not only the upper Bighorn but also the lower Bighorn are likely to Richmondian in age.
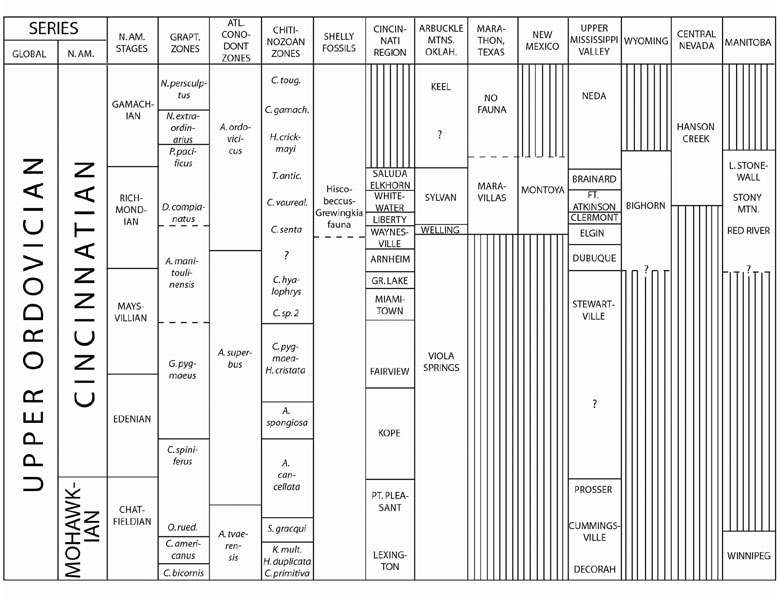
Figure 2. Correlation of some Cincinnatian formations in central and western North America. Note the large pre–Richmondian stratigraphic gap in several regions.
In Utah and eastern Nevada, closely similar conodont faunas are known from the Ely Springs and Fish Haven Formations. From the base of the presumably equivalent Hanson Creek Formation in central Nevada, Ross et al. (1979) illustrated A. ordovicicus, and Sweet (2000) described A. ordovicicus Zone faunas through the same formation at several other localities from strata with graptolites of the D. ornatus, P. pacificus, and N. extraodinarius Zones, chitinozoans of the T. scabiosa and the four next younger latest Ordovician (Richmondian and Gamachian) Zones (Soufiane and Achab, 2000), and locally, representatives of the Hiscobeccus–Grewingkia fauna. The upper part of the Hanson Creek also has a good record of the Gamachian d13C isotopic excursion. There can be no doubt that these successions are of Richmondian and Gamachian age although the basal part of the Hanson Creek, which has graptolites of the D. ornatus Zone, has been interpreted to be as old as late Edenian in some recent papers (e.g., Sweet, 2000) which is not in agreement with conodont, chitinozoan, and graptolite evidence (Figure 2).
Conclusions
Conodonts, graptolites, chitinozoans, and shelly fossils indicate that the Cincinnatian carbonate succession with its Hiscobeccus–Grewingkia shelly fauna is of Richmondian, and in some cases, Gamachian, age. This conclusion is in agreement with opinions expressed half a century (Twenhofel et al., 1954) or more ago, but it differs markedly from that of some recent workers (e.g., Sweet, 1979, 2000), who have proposed that the lower part of this succession is of late Edenian and Maysvillian age. The Richmondian–Gamachian carbonate successions in the Western Interior and Great Basin appear to represent a blanket–type deposit laid down in tropical shallow–water environments. Interestingly, these huge carbonate deposits are at least partly coeval with, but faunally mostly quite different from, the widespread Late Ordovician, pre–Hirnantian, carbonate interval in many regions round the Mediterranean (Bergström and Massa, 1992; Ferretti and Serpagli, 1999).
References
Achab, A. 1989. Ordovician chitinozoan zonation of Quebec and western Newfoundland. Journal of Paleontology, 63:14–24.
Bergström, S.M. 1971. Conodont biostratigraphy of the Middle and Upper Ordovician of Europe and eastern North America. Geological Society of America Memoir, 127:83–161.
Bergström, S.M. and Massa, D. 1992. Stratigraphic and biogeographic significance of Upper Ordovician conodonts from northwestern Libya. In: Salem, M.J., Hammuda, O.S., and Eliagoubi, B.A. (Eds.), The Geology of Libya, IV: 1323:1342. Elsevier Science Publishers.
Bergström, S.M. and Mitchell, C.E. 1986, The graptolite correlation of the North American Upper Ordovician standard. Lethaia, 19, 247–266.
Ferretti, A. and Serpagli, E. 1999. Late Ordovician conodont faunas from southern Sardinia, Italy: Biostratigraphic and paleogeographic implications. Bollettino della Società Paleontologica Italiana, 37:215–236.
Finney, S.C., Berry, W.B.N., Copper, J.D,. Ripperdam, R.L., Sweet, W.C., Jacobson, S. R., Soufiane, A., Achab, A., and Noble, P.J. 1999, Late Ordovician mass extinction: A new perspective from stratigraphic sections in central Nevada. Geology, 27:2115–218.
Goldman, D. and Bergström, S.M. 1997, Late Ordovician graptolites from the North American Midcontinent. Palaeontology, 40:965–1010.
Goldman, D., Bergsröm, S.M., and Mitchell, C.E. 1995, Revision of Zone 13 graptolite biostratigraphy in the Marathon, Texas standard succession and its bearing on Upper Ordovician graptolite biogeography. Lethaia, 28:115–128.
Howe, H.J. 1959. Montoya Group stratigraphy (Ordovician) of Trans–Pecos Texas. American Association Petroleum Geologists Bulletin, 43:2285–2332.
MacKenzie, P. and Bergström, S.M. 1993. Discovery of the zonal index fossil Amorphognathus ordovicicus in the Richmondian of Indiana: Implications for the regional correlation of the North American Upper Ordovician Standard. Geological Society of America, Abstracts with Programs, 25:A472.
Macomber, R.W. 1970. Articulate brachiopods from the upper Bighorn Formation (Late Ordovician) of Wyoming. Journal of Paleontology, 44:416–450.
Ross, R.J., Jr., Nolan, T.B., and Harris, A.G. 1980. The Upper Ordovician and Silurian Hanson Creek Formation of Central Nevada. U.S. Geological Survey Professional Paper, 1126C:C1–C22.
Soufiane, A. and Achab, A. 2000, Upper Ordovician and Lower Silurian chitinozoans from central Nevada and Arctic Canada. Review of Palaeobotany and Palynology, 113:165–187.
Sweet, W.C. 1979, Late Ordovician conodonts and biostratigraphy of the western Midcontinent Province. Brigham Young University Geology Studies, 26:45–86.
Sweet, W.C. 1984, Graphic correlation of upper Middle and Upper Ordovician rocks, North American Midcontinent Province, U.S.A. Palaeontological Contributions of the University of Oslo, 295:23–35.
Sweet, W.C. 2000. Conodonts and biostratigraphy of Upper Ordovician strata along a shelf to basin transect in central Nevada. Journal of Paleontology, 74:1148–1160.
Twenhofel, W.H., Bridge, J., Cloud, P.E., Cooper, B.N., Cooper, G.A., Cumings, E.R., Cullison, J.S., Kay, M., Liberty, B.A., McFarlan, A.C., Rodgers, J., Whittington, H.B., Wilson, A.E. and Wilson, C.W. 1954. Correlation of the Ordovician formations of North America. Geological Society of America Bulletin, 65:247–298.
Received: February 15, 2003
Accepted: June 15, 2003