The Niquivil Section, Precordillera of San Juan, Argentina, proposed gssp for the Lower/Middle Ordovician boundary
Guillermo L. Albanesi1, Marcelo G. Carrera1, Fernando L. Cañas2 and Matthew Saltzman3
1 CONICET – Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Casilla de Correo 1598, 5000 Córdoba, Argentina. E–mail: galbanesi@arnet.com.ar, mcarrera@com.uncor.edu
2 CONICET – Departamento de Geología, Universidad Nacional de Río Cuarto, Ruta Nacional 36 km 601, 5800 Río Cuarto, Argentina. E–mail: fcanas@exa.unrc.edu.ar
3 Department of Geological Sciences, The Ohio State University, Columbus, USA. E–mail: saltzman.11@osu.edu
Key words: Stratotype Section. Middle Ordovician. San Juan Formation. Precordillera. Argentina.
Introduction
Since the International Subcommission on Ordovician Stratigraphy (SOS) adopted the first appearance datum (FAD) of the conodont Tripodus laevis to mark the base of the Middle Ordovician Series (Webby, 1998), diverse problems arose upon the selection of this biostratigraphic marker. The Whiterock Narrows Section at Monitor Range, Nevada, proposed by Finney and Ethington (2000) as GSSP has been found to be of questionable usefulness in several respects. Detailed analyses of problems concerning both the selected conodont key species and the proposed section were posted on the "Ordovician Stratigraphy Discussion Forum" internet site (http://seis.natsci.csulb.edu/ordstrat1/default.htm).
The position of the Lower/Middle Ordovician Series boundary in the Argentine Precordillera was recently determined by means of conodont biostratigraphy to be the FAD of Tripodus laevis s.l. in the middle part of the San Juan Formation, Yanso Section, Potrerillo Mountain (Albanesi et al., 1998). In search of this horizon in graptolite facies, Ortega and Albanesi (1999) investigated new sections in northern Precordillera, which resulted in the recognition of the lower (but not lowest) Middle Ordovician biozones based on the Isograptus complex (beginning with I. victoriae maximus Zone), which begins at the T. laevis Zone (Albanesi et al., 1999). Graptolite biohorizons that lie within the critical interval we are considering were still not found in the Precordillera. After examination of diverse alternatives, Albanesi and Carrera (2001, and referred internet address) proposed the Niquivil Section of Central Precordillera as GSSP for the base of the Middle Ordovician by the FAD of the conodonts Texania heligma Pohler and Protoprioniodus aranda Cooper. This particular level was selected for being located just above the widely recognized Oepikodus evae Zone (referred taxa are considered key substitutes after the absence of T. laevis sensu stricto in the Precordillera).
The Niquivil Section
Location and stratigraphic framework
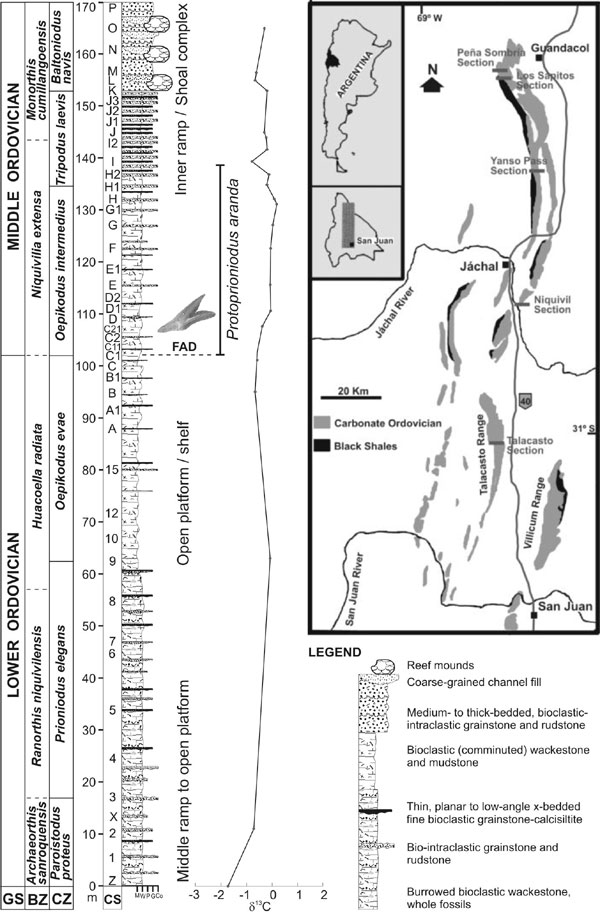
The Niquivil Section is located in the La Silla Range, which represents the easternmost part of the carbonate Eastern Tectofacies (Astini, 1992) of the Central Precordillera, San Juan Province. It is situated in Niquivil Village, on public land in town, along National Road 40, 22 km south of Jáchal City (Figure 1). The exposure of the San Juan Formation represents a thick northwest–dipping succession of carbonate rocks. The outcrops correspond to the lower and middle parts of the San Juan Formation (Keller et al., 1994; Cañas, 1999), including the Archaeorthis sanroquensis Zone at the base and the complete Monorthis cumillangoensis Zone at the top (brachiopod biostratigraphy sensu Benedetto, in press). Recently, Vaccari (2001) described new trilobite species from these strata.
The Lower/Middle Ordovician boundary interval occurs approximately at the boundary between the Huacoella radiata and Niquivilia extensa brachiopod zones without apparent lithological change. An important facies change is recorded at the boundary between the N. extensa and the Monorthis cumillangoensis zones. Fifty meters of pelmatozoan–rich grainstones and rudstones, including stromatoporoid–like organisms and small patch reefs occur in the basal part of the M. cumillangoensis Zone, which represents a sequence boundary after the sea level drop that follows the Oepikodus evae Zone transgression. Quaternary sediments cover the sequence, but it is precisely correlated with the nearest La Silla and Cerro Viejo sections. Both localities include the lower part of the formation (Paroistodus proteus Zone) (Lehnert, 1995). The Cerro Viejo Section includes the complete upper part of the formation (Ahtiella argentina Zone) up to the Lenodus variabilis Zone. The base of the overlying Los Azules Formation represents the Eoplacognathus suecicus and Undulograptus austrodentatus zones (Ottone et al., 1999).
Lithology and facies
The San Juan Limestone exhibits a wide range of carbonate textures, most of them bioclastic and/or skeletal (whole fossils), with variable amounts of lime mud, and is variably dolomitized. Marked pseudo–bedding distinguishes these rocks, showing alternating gray, massive horizons, and wavy to unevenly bedded levels that weather out as rubble, with a yellowish to tan cast where dolomite and argillaceous material are concentrated in solution seams and compacted burrows. Lithologies of the San Juan Formation were described in detail by Cañas (1999), who grouped the lithofacies in the unit into five associations, all representing fully–marine, open platform deposits, three of which occur in the Niquivil Section. These shallow–water carbonates are tectonically juxtaposed with slope and deep–basinal rocks to the west (Western Tectofacies of Astini, 1992), whereas by late Arenig time they graded northwards into deeper, outer ramp to intrashelf–basinal deposits (Figure 1).
Paleoecology
An integrated analysis of biofacies of the San Juan Formation was recently carried out by various authors (e.g., Carrera, 2001). According to these studies, the Niquivil Section includes the Archaeorthis biofacies in the lower part of the section, the Leptellinid biofacies, which includes the Ibexian/Whiterockian boundary as recognized in the Precordillera (FAD of T. laevis s.l. in Figures 1–2; cf., Ross and Ethington, 1991, Finney and Ethington, 2000), and the stromatoporoid biofacies in the uppermost part of the section. Cech and Carrera (2002) studied in detail the community replacement and distribution in the Niquivil Section. However, ongoing analyses revealed changes in community structure through the proposed Lower/Middle Ordovician boundary (sample NC1, Figures 1–2). Community replacement in the critical interval is not associated with important environmental change and it has been considered a modification of community type. In the uppermost Lower Ordovician levels communities are dominated by brachiopods. Biovolumetric values of the whole association show a diversified and biologically accommodated community. In the uppermost Lower Ordovician levels communities are dominated by brachiopods. Biovolumetric values of the whole association show a diversified and biologically accommodated community. In lowest Middle Ordovician levels communities are poorly diversified, including brachiopods, trilobites, laminar bryozoans, and dominated by gastropods (mainly macluritaceans).
Figure 1. Location map of Niquivil and other studied sections of Argentine Precordillera, and stratigraphic column of the Niquivil Section. CS: conodont samples, GS: Ordovician global series, BZ: brachiopod zones, CZ: conodont zones.
Conodont biofacial studies through the critical Lower/Middle Ordovician boundary in the San Juan Formation at the Yanso Section, Central Precordillera, were carried out by Albanesi (1998). The Argentine Precordillera is a peculiar area, where mixed cold– and warm–water faunas occur in the critical boundary interval. A turnover interval in the conodont faunas occurs close to the reference boundary, which ends the first cycle of conodont evolution (late Ibexian demise), with the extinction of several significant taxa.
Conodont record
The first conodont biostratigraphy of the Niquivil Section was published by Lehnert (1993), who defined six assemblage biozones. The present study presents a refined biostratigraphy for the referred critical interval, and follows the scheme established by Albanesi et al. (1998) for the Argentine Precordillera. It is based on 38 carbonate conodont samples, 3 kg each on average, which total ca. 120 kg. Samples were completely digested by conventional acid etching techniques (10% acetic acid) and yielded 5780 well–preserved conodonts that represent 54 multielement species (Figure 2). Samples were taken throughout a measured 170 m thick stratigraphic section, with 0.5 m sampling intervals within the critical boundary interval, where wackestones produced 150–200 elements per kg. Grainstones or coarser samples from the patch reefs–shoal interval that ends up the section produced less than 5 specimens per kg. Conodont elements show a Conodont Alteration Index of only 1.5–2 (Epstein et al., 1977), demonstrating low overburden paleotemperatures that allow for particular geochemical and paleomagnetic studies.
The FAD of Protoprioniodus aranda Cooper is selected as the key biohorizon for the global Lower/Middle Ordovician Series. It occurs in sample NC1, at the base of the Oepikodus intermedius Zone; i.e., 102 m above the base of the section, or 48.3 m below the basal level (NK) of the patch reefs–shoal interval that occurs at the top of the section. Among particular attributes as a key species, P. aranda has a cosmopolitan distribution, and occurs in virtually all environments; from shallow– to deep–, warm– to cold–water biofacies; e.g., Argentine Precordillera, Australia, Baltoscandia, South China, and North America (see references compiled by Albanesi in "Addenda" at http://seis.natsci.csulb.edu/ordstrat1/default.htm).
Another significant record at Niquivil Section is the presence of Protoprioniodus simplicissimus McTavish, which is considered the ancestral form of P. aranda according to a recent interpretation by Johnston and Barnes (1999). It is also interesting to observe that the FAD of P. aranda is part of the array to which the line of correlation has been fit for the graphic correlation of the upper part of the Ibex Area composite section, Millard County, Nevada (Sweet and Tolbert, in E. Taylor ed., 1997).
The vertical distribution of P. aranda is well–documented, it ranges from the uppermost levels of the North American Ibexian Series to the lowermost part of the Whiterockian Series; i.e., upper Reutterodus andinus through lower T. laevis zones (Ross et al., in E. Taylor ed., 1997). In Baltoscandia, P. aranda occurs in the upper part of the classic Oepikodus evae Zone, or in the Trapezognathus diprion Zone of the scheme of Bagnoli and Stouge (1997), where the index species O. evae is already absent; i.e., uppermost Latorpian Stage of the Oeland Series, which correlates with the upper Whitlandian Stage of the British Arenig Series (Fortey et al., 2000). The type stratum of P. aranda in the Amadeus Basin, Central Australia, was roughly correlated with the O. evae and Baltoniodus triangularis – B. navis zones (Cooper, 1981). In South China, P. aranda is recorded from upper O. evae Zone to lower B. triangularis Zone of the Dawan Formation in a locality close to the Huanghuachang Section (Wang Xiaofeng, epist. com., 2003). In the Argentine Precordillera, P. aranda has been reported from the diachronous top levels of the San Juan Formation and Lower Member of the Gualcamayo Formation in northern sections (Albanesi et al., 1999), and the middle part of the San Juan Formation in central sections, such as those at Yanso and Talacasto (Albanesi, unpubl. coll.).
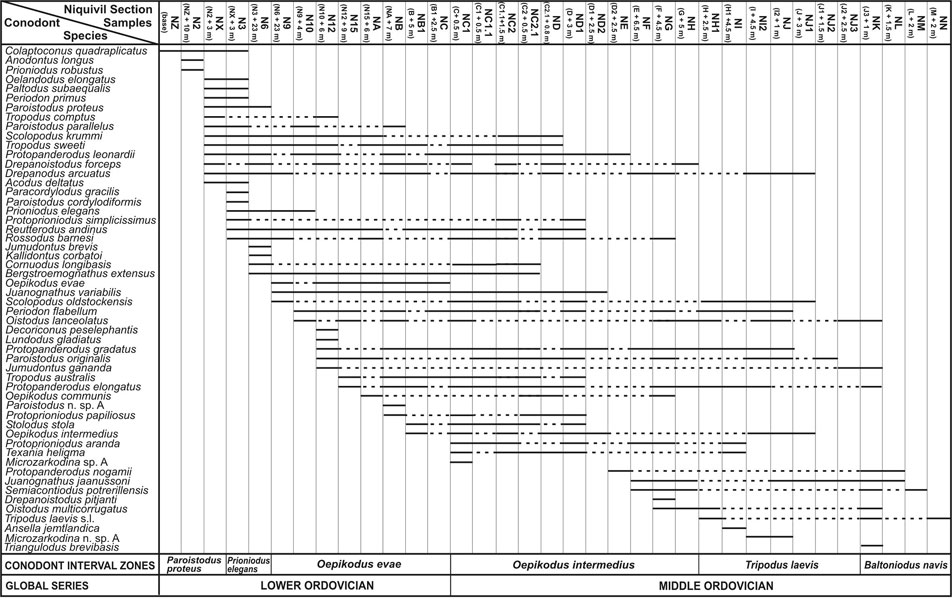
Figure 2. Conodont ranges and biozones of the Niquivil Section, Argentine Precordillera (no scale, only productive samples are registered).
The first appearance of P. aranda together with T. heligma occurs within the Isograptus victoriae lunatus Zone in upper slope sequences of western Newfoundland (Johnston and Barnes, 1999); i.e., in strata coeval with the Castlemainian 1 Stage of Australia. However, it occurs at lower Chewtonian (Ch 2) levels in some sections of Australia (A. VandenBerg, epist. com., 2002). The advantage of selecting P. aranda compared with other species is its wide paleogeographical and environmental distribution, on the contrary to other regional index, such as T. laevis or B. triangularis, which are apparently constrained to warm–, or cold–water, domains respectively, or restricted to particular facies. It is to note that for different regions and facies some short–ranging species (T. laevis, B. triangularis, T. heligma, T. diprion, and Microzarkodina spp.) appear close to the FAD of P. aranda, hence supporting an approximation of the proposed GSSP biohorizon (Figure 2).
Stable isotopes
The Carbon isotope values in the Niquivil Section record a shift from an Ibexian low of –2 per mil at the base of the sampled section (sample NZ) to a peak of just above 0 that occurs through an interval from ten to twenty meters above the proposed Lower/Middle Ordovician boundary (though without any significant excursion). Values from La Silla are similar to those from Niquivil and suggest the recorded trends are regional in scope. These trends in the Argentina sections are consistent with the carbon isotope stratigraphy obtained across the Ibexian/Whiterockian boundary at a well–studied section near Shingle Pass, Nevada, in which most analyzed samples fall between –1 and 0 per mil. In addition, if these trends are shown to be of global significance, it suggests that the burial ratio of organic carbon to carbonate carbon is likely to have increased during this time period (Figure 1).
Radiometric dating
New K–bentonites samples, interbedded with carbonate strata yielding the lowest records of P. aranda, whose heavy mineral suite proved suitable for radiometric dating, were recovered from the Talacasto Section. They yielded a mean of 469.8 ± 2.3 Ma (U–Pb SHRIMP dating by Baldo et al., this volume).
Conclusions
All attributes required for a GSSP by the International Commission on Stratigraphy (IUGS) are presented by the Niquivil Section; i.e., continuity of sedimentation and biozonation, completeness of exposures and adequate thickness without structural and metamorphic complication, abundance and variety of well–preserved fossils, favorable facies and correlation, amenability to isotopic age determination and magnetostratigraphy, accessibility, and preservation.
Acknowledgements
The authors acknowledge continued support by CONICET, and a special grant by SOS (ICS–IUGS) for present project. Drs. G. Ortega and S. H. Peralta assisted in field work. Drs. S.M. Bergström and G. Bagnoli reviewed the manuscript providing helpful suggestions.
References
Albanesi, G.L. 1998. Biofacies de conodontes de las secuencias ordovícicas del cerro Potrerillo, Precordillera Central de San Juan, R. Argentina. Actas XII Academia Nacional de Ciencias, Córdoba: 75–98.
Albanesi, G.L. and Carrera, M.G. 2001. Niquivil Section of Central Precordillera, Argentina, proposed as the Global Stratotype for the base of the Middle Ordovician Series. Geological Society of America Annual Meeting, Boston, Abstracts: A446.
Albanesi, G., Hünicken, M. and Barnes, C.R. 1998. Bioestratigrafía de conodontes de las secuencias ordovícicas del Cerro Potrerillo, Precordillera central de San Juan, R. Argentina. Actas XII Academia Nacional de Ciencias, Córdoba: 7–72.
Albanesi, G.L., Ortega, G., Barres, C.R. and Hünicken, M.A. 1999. Conodont–graptolite biostratigraphy of the Gualcamayo Formation (Middle Ordovician) in the Gualcamayo–Guandacol rivers area, Argentina Precordillera. In: Kraft, P. and Fatka, O. (Eds.), Quo Vadis Ordovician? 8th International Symposium on the Ordovician System, Prague, Acta Univesitatis Carolinae – Geologica 43 (1/2): 45–48.
Astini, R.A., 1992, Tectofacies ordovícicas y evolución de la cuenca eopaleozoica de la Precordillera Argentina. Estudios Geológicos, 48: 315–427.
Bagnoli, G. and Stouge, S. 1997. Lower Ordovician (Billingenian–Kunda) conodont zonation and provinces based on sections from Horns Udde, north Öland, Sweden. Bollettino della Società Paleontologica Italiana, 35: 109–163.
Baldo, E.G., Fanning, C.M., Rapela, C.W., Pankhurst, R.J., Casquet, C. and Galindo, C. 2003. U–Pb shrimp dating of rhyolite volcanism in the Famatinian belt and k–bentonites in the Precordillera. (this volume).
Benedetto, J.L. 2002. The Ordovician brachiopod faunas of Argentina: Chronology and biostratigraphic succession. In: F.G. Aceñolaza (ed.), Aspects on the Ordovician System of Argentina. INSUGEO, Tucumán, Serie Correlación Geológica 16: 87–106.
Cañas, F.L. 1999. Facies and sequences of late Cambian–early Ordovician carbonates of the Argentina Precordillera: A stratigraphic comparation whit Laurentia platforms. In: Keppie D. and Ramos, V. (Eds.). Laurentia–Gondwana connections before Pangea: Geological Society of America Special Paper 336: 43–62.
Carrera, M.G. 2001. Análisis de la distribución y composición de las biofacies de la Formación San Juan (Ordivícico Temprano), Precordillera Argentina. Ameghiniana, 38: 169–184.
Cech, N. and Carrera, M.G. 2002. Dinámica de las comunidades arenigianas de la Formación San Juan (Ordovícico), precordillera argentina. Ameghiniana, 39: 21–40.
Cooper, B.J. 1981. Early Ordovician conodonts from the Horn Valley Siltstone, central Australia. Palaeontology, 24(1): 147–183.
Epstein, A.G., Epstein, J.B. and Harris, L.D. 1977. Conodont color alteration – An index to organic metamorphism. United States Geological Survey Professional Paper, 995: 1–27.
Finney, S.C. and Ethington, R.L. 2000. Whiterock Narrows Section, Monitor Range, Nevada, proposed as the Global Stratotype for the base of the Middle Ordovician Series. Geological Society of America Annual Meeting, Reno, Abstracts: A391.
Fortey, R.A., Harper, D.A.T., Ingham, J.K., Owen, A.W., Parkes, M.A., Rushton, A.W.A. and Woodcock, N.H. 2000. A revised correlation of Ordovician rocks in the British Isles. The Geological Society. Special Report, 24: 1–83.
Johnston, D.I. and Barnes, C.R. 1999. Early and middle Ordovician (Arenig) conodonts from St. Pauls Inlet and Martin Point, Cow Head Group, western Newfoundland, Canada; 1. Biostratigraphy and paleoecology. Geologica et Palaeontologica, 33: 21–70.
Keller, M., Cañas,F.L., Lehnert,O., and Vaccari, N.E. 1994. The Upper Cambrrian and Lower Ordovician of the Precordillera (Western Argentina): Some stratigraphic reconsiderations. Newsletter on Stratigraphy, 31 (2): 115–132.
Lehnert, O. 1993. Bioestratigrafía de los conodontes arenigianos de la Formación San Juan en la localidad de Niquivil (Precordillera sanjuanina, Argentina) y su correlación intercontinental. Revista Española de Paleontología, 8 (2): 153–164.
Lehnert, O. 1995. Geodynamic processes in the Ordovician of the Argentine Precordillera: New biostratigraphic contraints. In: Cooper, J., Droser, M.L. and Finney, S.C. (Eds.), Ordovician Odyssey, 7th International Symposium on the Ordovician System. The Pacific section Society for Sedimentary Geology (SEPM), Fullerton, Book 77: 75–79.
Ortega G. and G.L. Albanesi. 1999. Graptolite biostratigraphy of the Gualcamayo Formation (Middle Ordovician) at the Los Sapitos creek section, Argentina Precordillera. In: P. Kraft and O. Fatka (Eds.). Quo Vadis Ordovician? 8th International Symposium on the Ordovician System, Prague, Acta Univesitatis Carolinae – Geologica 43 (1/2): 49–52
Ottone, E.G., Albanesi, G., Ortega, G. and Holfeltz, G. 1999. Palynomorphs, conodonts and associated graptolites from the Ordovician Los Azules Formation, Central Precordillera, Argentina. Micropaleontology, 45 (3): 225–250.
Ross, R.J. Jr. and Ethington, R.L. 1991. Stratotype of Ordovician Whiterock Series. Palaios, 6: 156–173.
Taylor, M.E. (ed.). 1997. Early Paleozoic Biochronology of the Great Basin, Wastern United States. U. S. Geological Survey Professional Paper, 1579: 1–115.
Vaccari, N.E. 2001. Trilobites de la Formación San Juan (Ordovícico Inferior), Precordillera Argentina. Ameghiniana, 38: 331–347.
Webby, B.D. 1998. Steps toward a global standard for Ordovician stratigraphy. Newsletter Stratigraphy, 36: 1–33.
Received: February 15, 2003
Accepted: June 15, 2003