Ordovician
Bivalvia and Rostroconchia of Argentina: An Updated Synthesis
Teresa M. SÁNCHEZ1
1 CONICET. Cátedra de Paleontología, Escuela de Biología, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Av. Velez Sarsfield 299, 5000 Córdoba, Argentina. E-mail: tsanchez@com.uncor.edu
Abstract:
ORDOVICIAN BIVALVIA
AND ROSTROCONCHIA OF ARGENTINA:
AN UPDATED
SYNTHESIS.
Knowledge of Ordovician Argentine bivalves and
rostroconchs substantially increased in the past ten years. Temporal
distribution of bivalves embraces the entire Ordovician, from the Tremadocian to
the late Asghill (Hirnantian), while rostroconchs are restricted to the
Tremadocian-early Llanvirn interval. Of 27 bivalve genera so far recognized in
the Argentine basins, 16 are endemics. Such unusually high endemism is
particularly evident in the Arenig volvaniclastic succession of the Famatina
basin and in the early Caradoc of the Precordillera, which confirms the
importance of Gondwanan shelves and peri-Gondwanan arc-related volcanic islands
in the early bivalve diversification.
Resumen:
BIVALVIA
Y ROSTROCONCHIA
ORDOVÍCICOS
DE ARGENTINA:
UNA SÍNTESIS
ACTUALIZADA.
Recientes
hallazgos en rocas Ordovícicas del oeste de Argentina han incrementado el número
de bivalvos y rostroconchos conocidos. La distribución temporal de los bivalvos
abarca desde el Tremadociano hasta el Ashgiliano tardío (Hirnantiano), mientras
que los rostroconchos están limitados al intervalo Tremadociano-Llanvirniano
temprano. De los 27 géneros de bivalvos reconocidos, 16 están restringidos a
cuencas argentinas, lo que representa un elevado porcentaje de endemismo. Este
fenómeno es particularmente notorio en la sucesión del Arenigiano medio de la
cuenca de Famatina y en los niveles de edad Caradociana temprana de la
Precordillera, lo que corrobora la importancia de las plataformas Gondwánicas y
arcos de islas volcánicas peri-Gondwánicas en la diversificación temprana de
los bivalvos.
Key
words: Bivalvia. Rostroconchia. Ordovician. Argentina
Palabras
clave: Bivalvia.
Rostroconchia. Ordovicico. Argentina.
Introduction
Although
bivalves are relatively unfrequent in the Ordovician rocks of western Argentina,
most typical Ordovician orders are represented. The richest and most abundant
bivalve faunas come from the siliciclastic rocks exposed in the Cordillera
Oriental, the southern part of the larger Central Andean basin, and from the
volcanosedimentary successions of the Sierra de Famatina. Conversely, the early
Ordovician carbonate rocks of the Precordillera basin, rich in brachiopods,
trilobites and sponges, have yielded up to now only isolated specimens of
bivalves and rostroconchs. In view that in limestones other molluscs having
aragonitic shells, namely gastropods, are locally very abundant, it is
reasonable to assume that the bivalve paucity in carbonate rocks is linked to
biotic limiting factors rather than tafonomic conditions, like decalcification
processes. As Babin (1993) stated, the Ordovician radiation of bivalves mainly
occurs around the siliciclastic peri-Gondwanan shelves.
However, it should be noted that volcanic phenomena may also have enhanced bivalve diversification, as occurs in the Famatina Range, where bivalves constitute a significant part of the nearshore biota (Waisfeld et al., 2001).
The
well-documented radiation underwent by most invertebrate clades during the
Ordovician is also verified among bivalves. In Argentina, several early groups
of bivalves, such as the Palaeotaxodonta (malletids, tironuculids,
praenuculids), Heteroconchia (cycloconchids, redonids), and Pteriomorphia
(colpomyids, mytiloids, ambonychiids, arcoidan) are well represented since the
Tremadocian and Arenig.
Although
rostroconchs have been documented in the three main basins of Argentina, they
never attain the diversity of other regions (e.g. Australia). So far, a total of
four genera and six species have been described. The record of Argentine
rostroconchs ranges from the late Cambrian (Ribeiria francae Sánchez,
2000) to the early Llanvirn (Sánchez, 1998). No younger species are yet known.
In spite of their scarcity, the four known genera represent the two orders of
the class recognized by Pojeta (1978), Ribeiroida and Conocardioida. The former
includes the genera Ribeiria Sharpe and Tolmachovia Howell and
Kobayashi, and the latter Eopteria Billings and Talacastella Sánchez.
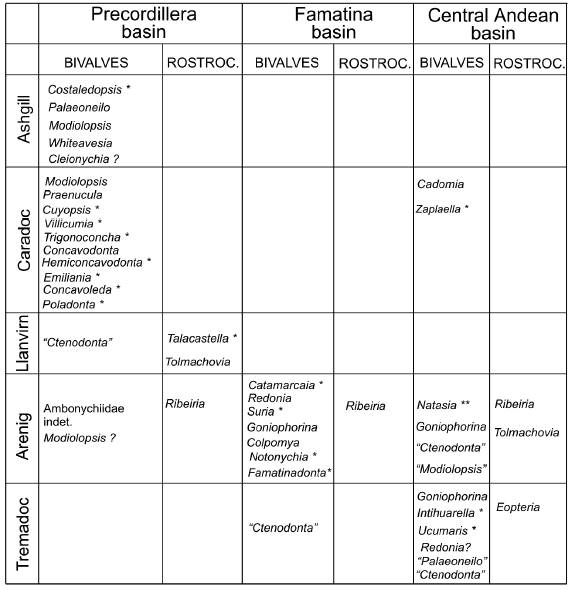
Figure 1. Spatial and temporal distribution of bivalve and rostroconch genera of Argentine Ordovician. Single asteriscs mark endemic genera, double asterisc indicates endemic genus of the Central Andean Basin (Argentina and Bolivia).
The
geographic distribution and stratigraphic ranges of Argentine bivalves and
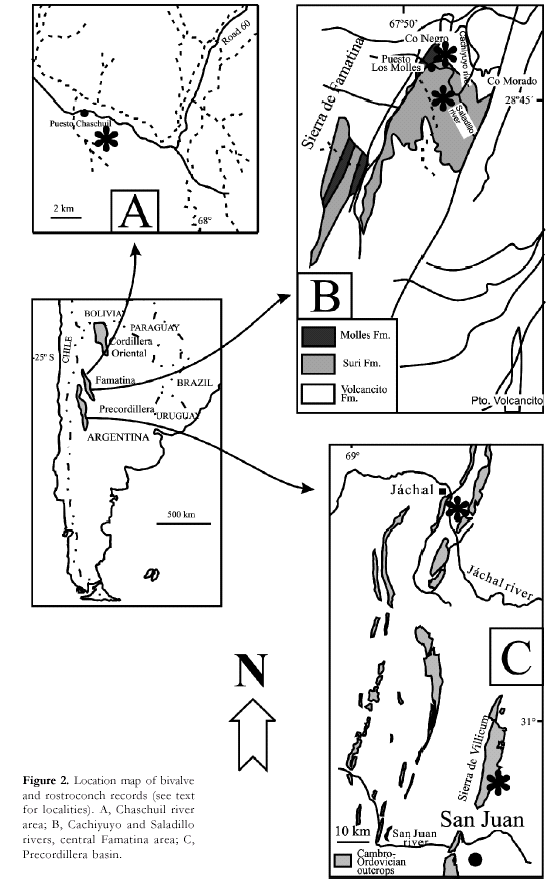
rostroconchs are shown in the figure 1. Both groups have been documented in the
three Ordovician basins of Argentina, the Precordillera terrane (Mendoza, San
Juan and La Rioja provinces), the arc-related Famatina basin (La Rioja and
Catamarca provinces) and the Central Andean Basin (Salta and Jujuy provinces)
(Figs. 2, 3). In the Precordillera basin, rostroconchs are confined to the
Arenig and early Llanvirn carbonate platform deposits, while bivalves, even
though sporadically present in these rocks, become relatively abundant in the
overlying Caradoc and Ashgill siliciclastic units in both deep and shallow water
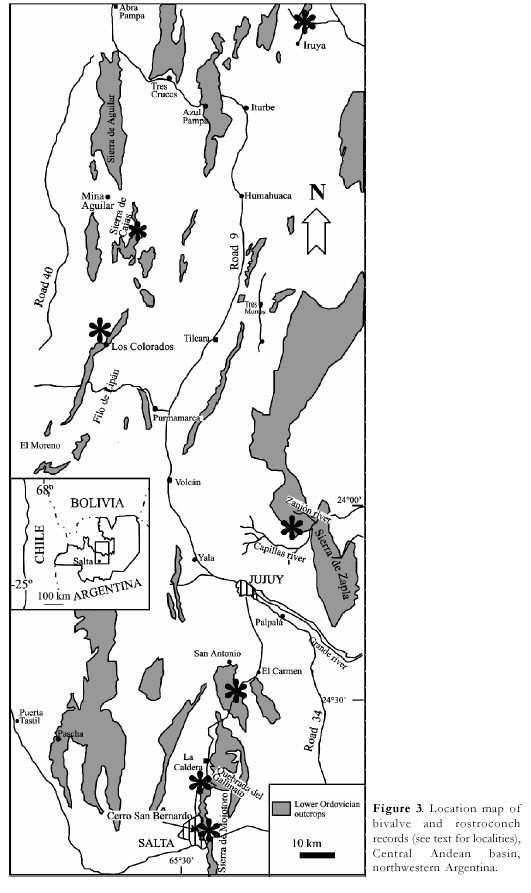
facies (Fig.2, C). In the Central Andean basin (Cordillera Oriental and Sierras
Subandinas) rostroconchs range from the latest Cambrian/earliest Tremadocian to
the mid-Arenig.
It
is interesting to note that the earliest known bivalves of Argentina, of
mid-Tremadocian age have been discovered in this basin. In general, bivalves are
a subordinate component of the Late Tremadoc and especially of the Arenig
communities of the Central Andean basin. Caradoc bivalves are locally abundant
in the Santa Gertrudis and Capillas formations which are exposed in the Sierra
de Mojotoro (Cordillera Oriental) and the Sierra de Zapla (Sierras Subandinas),
respectively (Fig.3). Finally, in the Sierra de Famatina, both bivalves and
rostroconchs are restricted to the Middle Arenig volcanosedimentary deposits
(Suri and Molles formations) cropping out in the Chaschuil area (Fig. 2 A) and
in the central region of the mountain belt, along the Cachiyuyo, Saladillo
Grande and Los Molles rivers) (Fig. 2 B).
A
list of bivalves and rostroconchs illustrated and described until now is given
in the figure 1. Bivalves which generic identification is in need of revision
include Ctenodonta laevigata Harrington, Ctenodonta minuscularia Harrington,
Ctenodonta famatinensis Harrington, Modiolopsis sanbernardica Harrington
(Harrington, 1938) from the Cerro San Bernardo (Acoite Formation), and Ctenodonta
bonetensis Aceñolaza, from Jagüé, La Rioja province (Aceñolaza, 1970).
From a biogeographical viewpoint, it is interesting to note that of these 27
bivalve genera, 16 are restricted to the Argentine basins, pointing out that an
important bivalve diversification occurred in the western Gondwana clastic
shelves, especially in the Famatina basin during the Arenig and in the
Precordillera basin during the early Caradoc.
There
are no bivalve genera in common among the three basins except Goniophorina, which
has been recorded from Famatina and the Cordillera Oriental. The remainder
genera are confined to a single basin. Goniophorina probably was an
opportunistic form capable to inhabit in both clastic shelves and volcanic
settings. With respect to the rostroconchs, Ribeiria has been documented
in the three basins in nearly contemporaneous horizons (middle Arenig),
represented by R.. spinosa in Famatina and Cordillera Oriental and by a R.
cf. compressa in the Precordillera. Tolmachovia, on the other
hand, appears in the Middle Arenig clastic successions of the Cordillera
Oriental and in slightly younger (early Llanvirn) carbonate rocks of the
Precordillera basin.
Stratigraphic
distribution of taxa
Tremadocian
Harrington
(1938) first noticed Tremadocian bivalves from the Cordillera Oriental (Iruya,
Jujuy Province) and from the Sierra de Famatina (Volcancito River, Catamarca
Province). From the former locality he reported Goniophorina
(Cosmogoniophorina) tenuicostata Harrington and Palaeoneilo iruyensis Harrington,
whereas from the ‘black shales of the Volcancito river’ (presently
Volcancito Formation) the only species recorded is Ctenodonta famatinensis Harrington.
Both the Iruya and Volcancito samples are early Tremadocian in age according to
the associated trilobites mentioned by Harrington (1938), the latter being
slightly older (Parabolina frequens argentina Zone). Because this
material represents the earliest known palaeotaxodonts it might play a
significant role in the debate about the origin of hinge types in bivalves.
Clearly, if these specimens are palaeotaxodonts, this strongly supports the
hypothesis that the taxodont pattern is ancestral to the actinodont dentition.
Unfortunatelly, hinges are not preserved neither in the specimens of ‘Palaeoneilo’
nor in those referred to ‘Ctenodonta’, so that in the available
matrial there is no evidence supporting its palaeotaxodont condition (for a
discussion on the hinge origin see Sánchez and Babin, 1998, and references
therein).
In
Argentina, the oldest known preserved dentitions are of actinodont type.
Specimens with this kind of dentition appear in the Floresta Formation exposures
near the Dique La Ciénaga (Sierra de Mojotoro, Jujuy Province) (Sánchez and
Vaccari, in press). According to the faunal content in other localities
(Moya et al., 1994; Moya, 1998) this unit has been referred to the Bienvillia
tegragonalis- Conophrys minutula Zone of Harrington and Leanza
(1957). Calibration of the trilobite zones with conodont and graptolite data
(Aceñolaza and Albanesi, 1997; Albanesi and Moya, 2000; Albanesi et al.,
2001; Moya et al., 1994; Ortega et al., 1997; Tortello and Rao,
2000) indicates that fossiliferous strata are late Tremadoc in age (P.
deltifer Zone). The bivalve assemblage from the Floresta Formation includes
the actinodont Intihuarella Sánchez, characterized by a very simple
actinodont type of hinge with a single posterior pseudolateral tooth. Another
associated actinodont-like form is Redonia? jujuyensis Sánchez,
in which the dentition is also reduced. The other two forms, Goniophorina
(Cosmogoniophorina) tenuicostata Harrington, and Ucumaris Sánchez,
are edentulous (Sánchez and Vaccari, in press).
Early
Tremadocian rostroconchs are represented by Eopteria agustini Sánchez (Sánchez,
2000) (Pl. 1, fig. 16), which comes from the Cardonal Formation exposed on the
eastern flank of the Sierra de Cajas, Jujuy province (Aceñolaza, 1968).
Arenig
Only
two bivalve species are presently known from the Precordillera, both from the
early Arenig Archaeorthis Zone (Herrera and Benedetto, 1991) of the San
Juan Formation: Modiolopsis? sp. and an undetermined ambonychiid (Sánchez,
2001a). Although this material is represented by a few specimens, its mention is
important as these species represent the oldest records of their respective
groups. The Middle Arenig beds of the same formation have yielded a single
species of rostroconch, Ribeiria cf. compressa Withfield (Sánchez,
1998).
In
the Cordillera Oriental the following early Arenig bivalves have been reported
by Harrington (1938) from the lower part of the Acoite Formation (San Bernardo
Formation sensu Moya, 1998):
Ctenodonta
laevigata Harrington, Ctenodonta minuscularia Harrington,
and Modiolopsis sanbernardica Harrington. Due to the absence of
diagnostic features (hinge, muscle scars) in the type material there is little
or no evidence to support these generic assignements. These species are in need
of revision.
Some
specimens from the Cerro San Bernardo assigned by Harrington (1938) to the
inarticulate brachiopod Obolus andinus Harrington correspond to the
rostroconch Tolmachovia, as was demonstrated by Gutierrez-Marco and Aceñolaza
(1991), who also reported Ribeiria spinosa Babin and Branisa from
slightly younger beds (Middle Arenig) cropping out at the Espinazo del Diablo,
east of Mina Aguilar, Jujuy province.
The
Acoite Formation contains shelly faunas dated as early to Middle Arenig on the
basis of graptolite content (Toro, 1994, 1997). It contains two bivalve species
(Sánchez, 1995b, 1997a, 1997b), Natasia boliviensis (Babin and Branisa)
(Pl.1, fig.1), and Goniophorina (Cosmogoniophorina) tenuicostata Harrington
(Pl. 1, fig. 2). The species boliviensis was first described from the
Arenig Sella Formation of Bolivia by Babin and Branisa (1987) and assigned to
the genus Ekaterodonta Babin. Additional information obtained from better
preserved and abundant specimens than those available at the time the Sella
species was described, including specimens of different size showing different
growth stages, revealed important differences with Ekaterodonta and
provided the basis for the erection of the genus Natasia Sánchez (Sánchez,
1995b) (Pl.1, fig. 1). The genera Natasia, Tironucula Morris and
Fortey, and Ekaterodonta all belong to the Family Tironuculidae Babin,
but there are important differences between them, especially in the growth
stages of dentition, justifying the recognition of two subfamilies, the
Tironuculinae, including Tironucula and Ekaterodonta, and
Natasiinae, including Natasia (Sánchez, 1997b).
Paleobiogeographically,
the record of Natasia boliviensis in the Sella and Acoite formations
constitute an additional evidence supporting the strong faunal links between the
Argentine and Bolivian parts of the Central Andean basin.
By
the Middle Arenig, an important diversification of bivalve faunas occurs in the
volcanic arcrelated Famatina basin. The Chaschuil river outcrops, in the
northern part of the basin, yielded excelently preserved bivalves (Aceñolaza
and Toselli, 1977; Sánchez and Babin, 1993, 1994), whereas the more continuous
successions exposed along the Cachiyuyo and Saladillo rivers (central Famatina)
have fournished both bivalves and rostroconchs (Sánchez and Babin, 1993, 1994;
Sánchez, 1997a).
In
the Chaschuil river area, the Loma del Kilómetro Member of the Suri Formation (sensu
Mángano and Buatois, 1996) contains the following species: Goniophorina
(Cosmogoniophorina) tenuicostata Harrington, Catamarcaia chaschuilensis (Aceñolaza
and Toselli) (Pl. 1, figs. 4 and 5), Suria ovalis Sánchez (Pl. 1, fig.
6), and Redonia suriensis Sánchez and Babin (Pl. 1, fig.7). Morphology
of Catamarcaia Sánchez and Babin is now known in considerable detail and
supports that this genus was the ancestor of the arcoidean clade (Sánchez,
1995a), which is in agreement with the phylogenetic ordering recently proposed
by Carter et al. (2000).
The
overlying Molles Formation, well exposed in the stratotype of the ‘Arroyo Los
Molles’ and partially in the Saladillo Grande river, has yielded the bivalves Redonia
riojana Sánchez, Notonychia emmae Sánchez, Famatinadonta
gonzaloi Sánchez (Pl. 1, fig.3), Colpomya elongata Sánchez (Pl. 1,
fig. 8), and Suria ovalis Sánchez (Sánchez, 2001b). The only
rostroconch recorded from this unit is Ribeiria spinosa Babin and
Branisa. In the Famatina volcanic-arc related basin bivalve diversification
mostly occurred in nearshore settings, the infaunal suspensivorous guild
occupied by Redonia, Famatinadonta, Catamarcaia and Colpomya
is dominant. Other guilds were occupied by single taxa: the epifaunal
suspensivorous by Notonychia and the infaunal detritivorous by Suria. The
guild occupation in the Central Andean basin differs in that the infaunal
suspensivorous guild was occupied by a single genus, Goniophorina (Waisfeld
et al., 2001).
Llanvirn-Caradoc
Sedimentary
rocks of Llanvirn age have only been documented in the Precordillera basin.
Abundant
external molds from the Cerro Condor Formation of Jagüé, La Rioja province,
(Aceñolaza and Bernasconi, 1969) were assigned to Ctenodonta bonetensis by
Aceñolaza (1970). However, the absence of preserved dentition in the available
material makes doubtful its assignement to the palaeotaxodonts. On the basis of
associated graptolites, the levels bearing C. bonetensis were referred to
the Llandeilo (Aceñolaza, 1970), and more recently Astini et al. (2000)
placed this unit within the Caradoc.
Talacastella
herrerai Sánchez (1986) (Pl. 1, fig. 15) and Tolmachovia sp.
(Sánchez, 1998) are the two known Llanvirn rostroconchs of Argentina. Both come
from the uppermost part of the San Juan Formation (early Llanvirn, Ahtiella Zone,
Herrera and Benedetto, 1991) of the Precordillera of San Juan.
Caradoc
By the Caradoc, the total number of bivalve genera increased considerably in the Precordillera basin (Sánchez, 1999a). At this time the bivalve diversification reaches a peak, probably related to a
global
increase of water temperature, also verified in other regions during the Caradoc
(Sánchez, 1999b).
Most
taxa were recorded in fossiliferous pebbles contained in the glacigenic
diamictite of the Don Braulio Formation (late Ashgill). Associated shelly fauna,
mainly brachiopods, indicate an early Caradoc age for these assemblages
(Benedetto, 1998). A striking feature is that such a diversification mostly
occurs within a single family, the praenuculids. It should also be noted that of
nine genera identified, seven are confined to the Precordillera, reflecting a
local radiation-extinction event (Sánchez, 1999b). A variable level of
endemism, though less marked than in bivalves, has also been verified in other
groups such as brachiopods, ostracods and trilobites (Benedetto et al.,
1999).
The
following praenuculids have been identified (Sánchez, 1999a): Praenucula sp.,
Cuyopsis symmetricus Sánchez (Pl. 1, fig. 12), Trigonoconcha acuta Sánchez
(Pl. 1, fig.10), Villicumia canteraensis Sánchez (Pl. 1, fig. 11), Concavodonta
ovalis Sánchez (Pl. 1, fig. 9), Hemiconcavodonta minuta Sánchez,
and Emiliania cuerdai Sánchez. Differences in orientation of the
chevroned teeth led to separate these forms into two subfamilies: Prenuculinae,
including Praenucula Pfab, Cuyopsis Sánchez, Trigonoconcha Sánchez,
Villicumia Sánchez, and Concavodontinae, including Concavodonta Babin
and Melou, Hemiconcavodonta Sánchez, and Emiliania Sánchez (Sánchez,
1999a). As Sánchez (1999b) suggested, the widespread Praenucula Pfab and
the perigondwanan Concavodonta Babin could be at the origin of the
subfamilies Praenuculinae and Concavodontinae, respectively.
Other
endemic taxa reported from this assemblage are Concavoleda braulense Sánchez
(Sánchez, 1999a) and the cycloconchid Poladonta sanjuanina Sánchez (Sánchez
et al., in press). An oustanding feature is that diversification occurs
almost entirely within the family Praenuculidae, which means that species
origination took place within a limited range of adaptive posibilities. Analysis
of modes of life of the seven endemic species of praenuculids indicates
infaunal, shallow burrowing habits, with similar degree of rocking movements.
Additionally, it is important to mention that apart from the cycloconchid Poladonta,
all the species of the association were detritus feeders, suggesting a muddy,
probably low-oxigenated bottom (Sánchez, 1999b).
Plate 1. Some Ordovician Bivalvia and Rostroconchia from Argentina. 1, Natasia bolviensis fragilis Sánchez, left valve, intermediate stage of dental development, early-Middle Arenig, Acoite Formation, Cordillera Oriental, CEGH-UNC 15082 (x 6). 2, Goniophorina (Cosmogoniophorina) tenuicostata Harrington, dorsolateral view, articulated specimen, early-Middle Arenig, Acoite Formation, Los Colorados locality, CEGH-UNC 15026, (x 2). 3, Famatinadonta gonzaloi Sánchez, right valve, Middle Arenig, Molles Formation, central Famatina, CEGH-UNC 18585 (x 3,7). 4, 5, Catamarcaia chaschuilensis (Aceñolaza and Toselli). 4, right valve, Middle Arenig, Suri Formation, Chaschuil river, CEGH-UNC 10530 (x 2,7). 5, latex replica of the same specimen, (x 3). 6, Suria ovalis Sánchez, laterodorsal view of left valve, umbo crushed to show dentition, Middle Ordovician, Suri Formation, Chaschuil and Cachiyuyo rivers, Famatina Range, CEGH- UNC 15016 (x 5,5). 7, Redonia suriensis Sánchez and Babin, right valve, Middle Ordovician, Suri Formation, Chaschuil river, holotype, CEGH-UNC 10474 (x 4). 8, Colpomya elongata Sánchez, right valve, Middle Arenig, Molles Formation, central Famatina, CEGH-UNC 18694 (x 3). 9, Concavodonta ovalis Sánchez, right valve, early Caradoc, basal diamictite of the Don Braulio Formation, Precordillera, CEGH-UNC 16169 (x 6). 10, Trigonoconcha acuta Sánchez, right valve, early Caradoc, basal diamictite of the Don Braulio Formation, Precordillera, CEGH-UNC 16167b (x 6). 11, Villicumia canteraensis Sánchez, incomplete left valve, early Caradoc, basal diamictite of the Don Braulio Formation, Precordillera, CEGH-UNC 17361 (x 6,5). 12, Cuyopsis symmetricus Sánchez, left valve, early Caradoc, basal diamictite of the Don Braulio Formation, Precordillera, holotype, CEGH-UNC 16719 (x 3,7). 13, Costaledopsis fuertensis Sánchez, articulated specimen, Ashgillian, N. persculptus Zone, unnamed unit, Cerro del Fuerte, Precordillera, holotype, CEGH-UNC 039 (x 2). 14, Modiolopsis cuyana Sánchez, left valve, articulated specimen, Ashgill, N. persculptus Zone, Don Braulio Formation, Villicum Range, Precordillera, holotype, CEGH-UNC 055 (x 2,7). 15, Talacastella herrerai Sánchez, right lateral view, early Llanvirn, San Juan Formation, Precordillera, CEGHUNC 17368 (x 4). 16, Eopteria agustini Sánchez, left lateral view, early Tremadocian, Cardonal Formation, Sierra de Cajas, holotype, CEGH-UNC 18465 (x 5). (Note: Fossils are housed in the Catedra de Estratigrafía y Geología Histórica, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, prefix CEGH-UNC).

The
almost coeval Las Plantas Formation exposed in the northern Precordillera
contains a single bivalve species, Modiolopsis inflata Sánchez (Sánchez,
1990).
The
Caradocian bivalves of the Central Andean basin are by far less diverse than in
the Precordillera.
The
Capillas Formation, cropping out in the Sierra de Zapla, and the Santa Gertrudis
Formation (Quebrada del Gallinato, Sierra de Mojotoro) have yielded Cadomia
typa De Tromelin and a cycloconchid previously assigned to Cycloconcha cf.
C. oblonga Foerste (Sanchez, 1986) but now assigned to Zaplaella
capillaensis Sánchez (Sánchez et al., in press). Cadomia typa is
a typical form of the Armoricain Massif and Iberia, and its record in the
Central Andean basin indicates a clear faunal link between northwestern
Argentina and these peri-Gondwanan areas. The age of this units has been matter
of debate.
Harrington
and Leanza (1957) referred this unit to the early Llanvirn, but Monaldi and
Monaldi (1978), also based on the trilobite content, suggested a younger early
Caradoc age. On the basis of the bivalve fauna Sánchez (1986) also supported a
Llandeilo age for the Santa Gertrudis Formation.
According
to Albanesi and Rao (1996), the conodont assemblage from the beds containing the
shelly fauna indicates the early Caradoc. The Capillas Formation, bearing a
similar bivalve and trilobite fauna, can be confidently correlated with the
Santa Gertrudis Formation (Waisfeld, 1996).
Ashgill
Ashgill
bivalves are presently known only from the Precordillera basin. The uppermost
part of the Trapiche Formation exposed at the Gualcamayo river section has
yielded a poorly preserved ambonychiid (Cleionychia? sp., Sánchez, 1990)
and other undeterminable forms. These levels have been considered as late
Ashgill on the basis of the brachiopod Trucizetina Benedetto (1999), even
though no typical elements of the Hirnantia Fauna have been discovered.
The Late Ashgill (Hirnantian) Don Braulio Formation contains Modiolopsis
cuyana Sánchez (Pl. 1, fig. 14) and Paleoneilo sp. (Sánchez, 1990).
Calcareous lenses developed at the first stages of the latest Ashgill
transgression yielded abundant specimens of Modiolopsis cuyana associated
with the brachiopods Hirnantia sagittifera (McCoy), Dalmanella
testudinaria (Dalman), and Paromalomena polonica (Temple) (Benedetto,
1986), the trilobite Dalmanitina sudamericana Baldis and Blasco (1975),
and encrusting bryozoa, all of which integrate the shallow-water Modiolopsis-Hirnantia
community. This relatively low diverse community probably reflects a
fluctuating resource supply in a cold-water shallow environment (Sánchez et
al., 1991).
The
unnamed latest Ashgill (N. persculptus Zone) mudstone succession exposed
in the Cerro del Fuerte area (Benedetto et al., 1986) has provided
specimens of Costaledopsis fuertensis Sánchez (Pl. 1, fig. 13) and Whiteavesia
sp. (Sánchez, 1990). This unit probably represents a deep-water equivalent
of the late Ashgill succession of the Sierra de Villicum.
Taxonomic
ordering of bivalve species
PALAEOTAXODONTA
-Family
Praenuculidae McAlester, 1969
Subfamily
Praenuculinae Sánchez, 1999a
Praenucula
sp.
Costaledopsis
fuertensis Sánchez, 1990
Cuyopsis
symmetricus Sánchez, 1999a
Trigonoconcha
acuta Sánchez,
1999a
Villicumia
canteraensis Sánchez,
1999a
Subfamily
Concavodontinae Sánchez, 1999a
Concavodonta
ovalis Sánchez,
1999a
Hemiconcavodonta
minuta Sánchez,
1999a
Emiliania
cuerdai Sánchez,
1999a
-
Family Malletiidae Adams and Adams, 1858
Paleoneilo
sp.
‘Palaeoneilo’
iruyensis Harrington, 1938
Concavoleda
braulense Sánchez,
1999a
Cadomia
typa De Tromelin,
1877
Suria
ovalis Sánchez, 1997a
-
Family Tironuculidae Babin, 1982
Subfamily
Natasiinae Sánchez, 1997b
Natasia
boliviensis (Babin
and Branisa, 1978)
-
Family Ctenodontidae Wöhrmann, 1893
‘Ctenodonta’
famatinensis Harrington, 1938
‘Ctenodonta’minuscularia
Harrington, 1938
‘Ctenodonta’laevigata
Harrington, 1938
‘Ctenodonta’
bonetensis Aceñolaza, 1970
HETEROCONCHIA
-
Family Cycloconchidae Ulrich, 1884
Famatinadonta
gonzaloi Sánchez,
2001b
Zaplaella
capillaensis Sánchez
(in Sánchez et al., in press)
Poladonta
sanjuanina Sánchez
(in Sánchez et al., in press)
-
Familly Intihuarellidae Sánchez (in Sánchez and Vaccari, in press)
Hintihuarella
simplicidentata Sánchez (in Sánchez and Vaccari, in
press)
-
Family Redoniidae Babin, 1966
Redonia
suriensis Sánchez and Babin, 1994
Redonia
riojana Sánchez,
1997a
Redonia
? jujuyensis Sánchez (in Sánchez and
Vaccari, in press)
PTERIOMORPHIA
-
Family Modiolopsidae Fischer, 1887
Modiolopsis
cuyana Sánchez,
1990
Modiolopsis
inflata Sánchez,
1990
‘Modiolopsis’
sanbernardica Harrington, 1938
Modiolopsis?
sp.
Whiteavesia
sp.
-
Family Colpomyidae Pojeta & Gilbert-Tomlinson, 1977
Colpomya
elongata Sánchez,
2001b
-
Family Ambonychiidae Miller, 1877
Cleionychia?
sp.
Notonychia
emmae Sánchez, 2001b
ARCOIDA
-
Family Catamarcaiidae Cope, 2000
Catamarcaia
chaschuilensis (Aceñolaza
and Toselli, 1977)
ANOMALODESMATA
-
Family Modiomorphidae Miller, 1877
Goniophorina
(Cosmogoniophorina) tenuicostata Harrington,
1938
-
Family Ucumariidae Sánchez (in Sánchez and Vaccari, in press)
Ucumaris
conradoi Sánchez (in Sánchez and Vaccari, in press)
Taxonomic
ordering of rostroconchian species
RIBEIROIDA
-
Family Ribeiridae Kobayashi, 1933
Ribeiria
spinosa Babin and Branisa, 1987
Ribeiria
cf. R. compressa Whitfield, 1886
-
Family Technophoridae Miller, 1889
Tolmachovia
andina (Harrington,
1938)
Tolmachovia
sp.
CONOCARDIOIDA
-
Family Eopteriidae Miller, 1889
Eopteria
agustini Sánchez,
2000
Talacastella
herrerai Sánchez,
1986
Acknowledgements.
I thank the volume editor, F. G. Aceñolaza, for the
invitation to participate in this volume.
Paleontologic
studies carried out in the last years were made possible through support of the
Consejo Nacional de Investigaciones Científicas y Técnicas, CONICET (Grant
05/1316), and the Agencia Nacional de Promoción Científica y Tecnológica,
ANPCyT (Grant 5387).
References
Aceñolaza,
F.G., 1968. Geología estratigráfica de la región de la Sierra de Cajas,
Depto. Humahuaca (Provincia de Jujuy). Revista de la Asociación Geológica
Argentina, 23: 207-222.
Aceñolaza,
F.G. 1970. Fósiles ordovícicos del área del Río Bonete, Depto. Jagüe,
provincia de La Rioja. Acta Geológica Lilloana, 10:313-328. Aceñolaza,
G.F., and Albanesi, G.L. 1997. Conodont-trilobite biostratigraphy of the Santa Rosita
Formation
(Tremadoc)
from Chucalezna, Cordillera Oriental, northern Argentina. Ameghiniana,
34: 113 Aceñolaza, F.G. and Bernasconi, A., 1969. Acerca del primer hallazgo de
una fauna ordovícica en el sector norte de la Precordillera riojana. Revista
de la Asociación Geológica Argentina, 24: 79-84.
Aceñolaza,
F.G., and Toselli, A., 1977. Observaciones geológicas y paleontológicas sobre el Ordovícico de
la zona de Chaschuil, Provincia de Catamarca. Acta Geológica Lilloana,
14: 55-81.
Albanesi,
G.L., and Moya, M.C. 2000. New stratigraphic section to define the
Cambrian-Ordovician boundary in Eastern Cordillera, northwestern Argentina. In
Aceñolaza, G.F. and Peralta, S. (eds.) Cambrian from the southern edge.
INSUGEO, Miscelanea 6: 114-116.
Albanesi,
G. and Rao, R., 1996. Conodont fauna from the Santa Gertrudis Formation
(middle-late Ordovician), eastern Cordillera, northwestern Argentina. Sixth
International Conodont Symposium (ECOS VI), Abstracts: 3. Warszawa.
Albanesi,
G.L., Ortega, G., and Zeballo, F. 2001. Late Tremadocian conodont-graptolite
biostratigraphy from NW Argentine basins. The Guide Book, Joint Field Meeting
IGCP 410/IGCP 421 in Mongolia, pp. 121-123.
Astini,
A.A., Brussa, E.D., and Mitchell, C.E., 2000. Revisión estratigráfica y consideraciones
paleogeográficas de la tectofacies occidental de la Precordillera argentina. Revista
de la Asociación Geológica Argentina, 55: 378-386.
Babin,
C., 1993. Rôle des plates-formes gondwaniennes dans les diversifications des
mollusques bivalves durant l’Ordovicien. Bulletin de la Societé géologique
de France, 164: 141-153.
Babin,
C., and Branisa, L., 1987. Ribeiria,
Peelerophon y otros moluscos del Ordovícico de Bolivia. IV Congreso
Latinoamericano de Paleontología, Santa Cruz, Bolivia, 1: 119-129.
Baldis,
B.A. and Blasco, G., 1975. Primeros trilobites ashgillianos del Ordovícico Sudamericano. Primer
Congreso Argentino de Paleontología y Bioestratigrafía, Actas 1: 33-48. Tucumán.
Benedetto,
J.L., 1998. Braquiópodos
caradocianos en los bloques de la diamictita glacigénica de la Formación Don
Braulio (Ashgilliano), Sierra de Villicum, Precordillera Argentina. Ameghiniana,
35: 393-412.
Benedetto,
J.L., 1999. Braquiópodos
de la Formación Trapiche, Ordovícico tardío de la Precordillera de La Rioja y
San Juan, Argentina. Ameghiniana, 36: 323-337.
Benedetto,
J.L., Ortega, G., Brussa, E., and Toro, B., 1986. Estratigrafía y fauna de la
secuencia Neo-Ordovícica y Llandoveriana del Río Escondido (flanco occidental
del Cerro del Fuerte), Provincia de San Juan. Primeras Jornadas sobre Geología
de la Precordillera, Actas 1: 128-133.
Benedetto,
J.L., Sánchez, T.M., Carrera, M.G., Brussa, E.D., and Salas. M.J., 1999.
Paleontological constraints on successive paleogeographic positions of
Precordillera terrane during the Early Paleozoic. In: Keppie, D. and
Ramos, V.A. (eds.) Laurentia-Gondwana connections before Pangea. Geological
Survey of America, Special Paper 336: 21-42.
Carter,
J.G., Campbell, D.C., and Campbell, M.R., 2000. Cladistic perspectives on early
bivalve evolution. In E.M. Harper, J.D. Taylor, and J.A. Crame (eds.) The
Evolutionary Biology of the Bivalvia. Geological Society, London, Special
Publications, 177: 47-79.
Harrington,
H.J., 1938. Sobre las
faunas del Ordoviciano inferior del Norte argentino. Revista del Museo de La
Plata (Nueva Serie), t. 1, Sección Paleontología, p. 109-289.
Harrington,
H.J. and Leanza, A.F., 1957. Ordovician trilobites of Argentina. Department
of Geology, University of Kansas, Special Publication 1, 276 p.
Herrera,
Z., and Benedetto, J.L., 1991. Early Ordovician brachiopod faunas of the
Precordillera basin, Western Argentina: Biostratigraphy and paleobiogeographical
affinities. In: MacKinnon, D.I., Lee, D.E., and Campbell, J.D. (eds.) Brachiopods
through time. p. 283-301. A.A. Balkema, Rotterdam.
Gutiérrez-Marco,
J.C., and Aceñolaza, F.G., 1991. Ribeiria and Tolmachovia (Mollusca,
Rostroconchia) en el Ordovícico inferior de la Cordillera Oriental Argentina. Zentralblatt
für Geologie und Paläontologie 1: 1799-1814.
Mángano,
M.G. and Buatois, L.A., 1996. Shallow marine event sedimentation in a volcanic
arc-related setting: The Ordovician Suri Formation, Famatina Range, northwest
Argentina. Sedimentary Geology, 105: 63-90.
Monaldi,
C.R. and Monaldi, O.H., 1978. Hallazgo de una fauna en la Formación Santa Gertrudis (Ordovícico),
provincia de Salta, República Argentina. Revista de la Asociación Geológica
Argentina, 33: 245-246.
Moya,
M.C., 1998. El Paleozoico inferior en la sierra de Mojotoro, Salta-Jujuy. Revista
de la Asociación Geológica Argentina, 53: 219-238.
Moya,
M.C., Malanca, S., Monteros, J.A., and Cuerda, A., 1994. Bioestratigrafía del
Ordovícico inferior en la Cordillera Oriental argentina, basada en graptolitos.
Revista Española de Paleontología, 9: 91-104.
Ortega.
G., Tortello, M.F., Rao, R.I., and Aceñolaza, G.F. 1997. Faunas del Ordovícico
Inferior en el área del Angosto de Lampazar, Cordillera Oriental, provincia de
Salta, Argentina. Ameghiniana, 34: 123.
Pojeta,
J.Jr., 1978. Class
Rostroconchia. In Boardman, R.S., Cheetham, A.H., and Rowell, A.J. (eds.)
Fossil Invertebrates. p. 358-380. Blackwell Scientific Publications, Palo
Alto, Oxford, London, Melbourne.
Sánchez,
T.M., 1986. Una fauna de bivalvos en la Formación Santa Gertrudis (Ordovícico)
de la Provincia de Salta (Argentina). Ameghiniana, 23: 131-139.
Sánchez,
T.M., 1990. Bivalvos del Ordovícico medio-tardío de la Precordillera de San
Juan (Argentina). Ameghiniana,
27: 251-261.
Sánchez,
T.M., 1995a. Comments on the genus Catamarcaia Sánchez & Babin and
the origin of the Arcoida. Geobios,
28: 343-346.
Sánchez,
T.M., 1995b. Un nuevo género de Tironuculidae (Bivalvia, Palaeotaxodonta) en el
Arenigiano del Noroeste Argentino. Geobios, 28: 683-690.
Sánchez,
T.M., 1997a. Additional
mollusca (Bivalvia and Rostroconchia) from the Suri Formation, early Ordovician
(Arenig), western Argentina. Journal of Paleontology, 71: 1046-1054.
Sánchez,
T.M., 1997b. Natasiinae,
a new subfamily of Tironuculidae (Bivalvia, Palaeotaxodonta) from the Acoite
Formation (early-middle Arenig), northwestern Argentine. Geobios,
M.S. nº 20: 471-475.
Sánchez,
T.M., 1998. Rostroconchia (Mollusca, Diasoma) en la Formación San Juan (Ordovícico
temprano), Precordillera Argentina. Ameghiniana, Nota Paleontológica,
35:345-347.
Sánchez,
T.M., 1999a. New
late Ordovician (early Caradoc) bivalves from the Sierra de Villicum (Argentine
Precordillera). Journal of Paleontology, 73: 66-76.
Sánchez,
T.M., 1999b. Caradoc
bivalves from the Argentine Precordillera: a local radiation-extinction event. Geobios,
32: 333-340.
Sánchez,
T.M., 2000. The
oldest record of Rostroconchia (Mollusca) from western Gondwana (Cordillera
Oriental, northwestern Argentina). Ameghiniana,
37: 265-269.
Sánchez,
T.M., 2001a. Bivalvia (Mollusca) de la Formación San Juan (Ordovícico
temprano, Precordillera Argentina). Ameghiniana, 38: 471-475.
Sánchez,
T.M., 2001b. Moluscos bivalvos de la Formación Molles (Arenigiano medio),
sierra de Famatina. Ameghiniana,
38: 185-193.
Sánchez,
T.M. et Babin, C., 1993. Un
insolite mollusque bivalve, Catamarcaia n. g. de l’Arenig (Ordovicien
inférieur) d’Argentine. Comptes Rendus, Academie des Sciences, Paris.
316, Série II: 265-271.
Sánchez,
T. M. y Babin, C. 1994. Los géneros Redonia y Catamarcaia (Mollusca,
Bivalvia) de la Formación Suri (Ordovícico temprano, Oeste de Argentina) y su
interés paleobiogeográfico. Revista Española de Paleontología,
9:81-90.
Sánchez,
T.M. and Babin, C., 1998. The origin of actinodont from taxodont dentitions or
vice-versa: An unnecessary controversy?. In: P.A.Johnston and J. W.
Haggart (eds.) Bivalves: an eon of evolution. University of Calgary
Press, pp.409-412.
Sánchez,
T.M., Benedetto, J.L., and Brussa, E., 1991. Late Ordovician stratigraphy,
paleoecology, and sea level changes in the Argentine Precordillera. In: Barnes,
C.R., and Williams, S.H, (eds.) Advances in Ordovician Geology.
Geological Survey of Canada, Paper 90-9: 245-258.
Sánchez,
T.M., and Vaccari, N.E., in press. Ucumariidae new family (Bivalvia,
Anomalodesmata) and other bivalves from the Early Ordovician (Tremadoc) of
northwestern Argentina. Ameghiniana.
Sánchez,
T.M., Marengo, L., and Astini, R., in press. Late Ordovician cycloconchids
(Bivalvia, Heteroconchia) from western Argentina. Ameghiniana.
Toro,
B.A., 1994. Las zonas de Didymograptus (Didymograptellus) bifidus (Arenigiano
medio) y Didymograptus (Corymbograptus) deflexus (Arenigiano inferior) en
la Formación Acoite, Cordillera Oriental, Argentina. Ameghiniana, 31:
209-220.
Toro,
B.A., 1997. La fauna de graptolitos de la Formación Acoite, en el borde
occidental de la Cordillera Oriental Argentina. Análisis bioestratigráfico. Ameghiniana,
34: 393-412.
Tortello,
M.F., and Rao, R.I. 2000. Trilobites
y conodontes del Ordovícico temprano del Angosto de Lampazar (provincia de
Salta, Argentina). Boletín Geológico y Minero, 111:61-84.
Waisfeld,
B.G., 1996. Revisión
de la Zona de Hoekaspis schlagintweiti Harrington & Leanza, Ordovícico
del Noroeste de Argentina. Memorias del XII Congreso Geológico de Bolivia,
Tarija, p.915-921.
Waisfeld,
B.G., Sánchez, T.M., Benedetto, J.L., and Carrera, M.G., 2001. Arenig faunal
assemblages from western Argentina: biodiversification and paleogeographical
constraints. Early Palaeozoic Palaeogeographies and Palaeobiogeographies of
western Europe and north Africa. Abstracts, p. 73. Lille, september, 2001.
Recibido:
25 de Agosto de 2002
Aceptado: 10 de Octubre de 2002