Trilobites
from the Base of the Ordovician System in Northwestern Argentina
M.
Franco TORTELLO 1, Susana B. ESTEBAN2 and Guillermo F. ACEÑOLAZA2
1 Departamento Paleontología Invertebrados, Museo de La Plata, Paseo del Bosque s/n°, 1900 La Plata, Argentina. E-mail: tortello@museo.fcnym.unlp.edu.ar
2 Instituto Superior de Correlación Geológica (INSUGEO), Facultad de Cs. Naturales e Inst. Miguel Lillo, Universidad Nacional de Tucumán, Miguel Lillo 205, 4000 Tucumán, Argentina. E-mail: insugeo@unt.edu.ar
Abstract.-
TRILOBITES FROM THE BASE OF THE ORDOVICIAN
SYSTEM IN NORTHWESTERN ARGENTINA:
The Cambrian-Ordovician boundary interval is well
represented within the trilobite Parabolina (Neoparabolina) frequens
argentina Biozone in northwestern Argentina. The lower part of the biozone
is characterized by a diverse agnostoid-olenid assemblage assignable to the
latest Cambrian. On the other hand, the upper part of the unit is diagnosed by
the occurrence of Jujuyaspis keideli Kobayashi (early Tremadocian).
Stratigraphic information on seven classical sections from the base of the
Ordovician System in the Cordillera Oriental and Famatina Range is summarized.
The
Jujuyaspis keideli Subzone is formally described on the basis of the
stratotype of the río Volcancito section (upper part of the “Lower Member”
of the Volcancito Formation), where J. keideli is recorded in association
with early planktic graptolites. In addition, distinctive trilobites from the
overlying Kainella meridionalis Biozone are here reillustrated.
Resumen:
TRILOBITES
DE LA BASE DEL SISTEMA
ORDOVÍCICO
EN LA REGIÓN NOROESTE DE ARGENTINA:
El intervalo que
incluye la transición entre los Sistemas Cámbrico y Ordovícico se encuentra
bien representado en la región noroeste de Argentina, dentro de la Biozona de Parabolina
(N.) frequens argentina. El sector inferior de la biozona se caracteriza por
una diversa asociación de agnóstidos y olénidos asignable al Cámbrico tardío,
mientras que la parte superior se diagnostica con el registro de Jujuyaspis
keideli Kobayashi (Tremadociano temprano). En esta comunicación se resume
la información estratigráfica de siete secciones clásicas donde se encuentra
representada la base del Ordovícico en el Sistema de Famatina y Cordillera
Oriental. Asimismo se define formalmente la Subzona de Jujuyaspis keideli en
base a la asociación de J. keideli y Rhabdinopora flabelliformis en
la sección de río Volcancito (sector superior del “Miembro Inferior” de la
Formación Volcancito, Sistema de Famatina). Por último, se re-ilustran
trilobites que caracterizan la suprayacente Biozona de Kainella meridionalis.
Key
words: Trilobites. Early
Tremadocian. Cordillera Oriental. Famatina. Argentina.
Palabras
clave: Trilobites. Tremadociano temprano. Cordillera
Oriental. Famatina. Argentina.
Introduction
After
debating for 25 years, the International Working Group on the
Cambrian-Ordovician Boundary (COBWG) has expressed its preference for Green
Point (Cow Head Group, Green Point Formation, Western Newfoundland) as the
Global Stratotype Section and Point (GSSP) for base of the Ordovician System
(approved by ISOS, ICS and IUGS; Cooper et al., 2001). The Green Point
section (latest Cambrian-Arenig) is mainly composed of alternating shale,
limestone and grainstone representing a base-of-slope depositional environment.
The Cambrian-Ordovician boundary interval is characterized by a complete record
of conodonts and graptolites. The base of the Ordovician System is indicated by
the first appearance of the conodont Iapetognathus fluctivagus Cooper et
al., which lies 4.8 m below the FAD of planktic graptolites (Rhabdinopora
praeparabola Zone) (Cooper et al., 2001). Unfortunately,
shelly fossils are rare at Green Point. However, Fortey et al. (1982)
described trilobites from adjacent, more upslope sections and discussed their
biostratigraphic implications.
The
lowermost Tremadocian correlates with the FAD of the olenid trilobite Jujuyaspis
Kobayashi, a cosmopolitan genus originally described from Argentina. Many
typical sections spanning the Cambrian-Ordovician boundary are characterized by
conodonts, planktic graptolites, and endemic trilobites associated with Jujuyaspis
[e.g. Lawson Cove (western Utah, U.S.A), Yongwol (centraleastern Korea) -
Magezhuang (North-East China) and Naersnes (Oslo region, Norway)]. In North
America, Miller and Stitt (1995) correlated the bases of the Symphysurina
bulbosa Subzone and the Iapetognathus Zone with the base of the
Ordovician, pointing out the occurrence of Jujuyaspis in these units
(e.g. Norford, 1969; Winston and Nichols, 1967; Taylor and Repetski, 1995;
Westrop, 1986; Stitt and Miller, 1987; Dean, 1989; Miller and Taylor, 1995; Loch
et al., 1999). Besides, in North China and Korea the genus has been
described in association with Yosimuraspis in the homonymous zone (early
Ordovician) (e.g. Chen et al., 1980; Zhou and Zhang, 1985; Kim and Choi,
2000), whereas it occurs within the range of Cordylodus lindstromi,
Iapetognathus and Rhabdinopora flabelliformis socialis in Norway,
Colombia, Bolivia and Argentina (Kobayashi, 1936; Harrington and Leanza, 1957;
Henningsmoen, 1957; Pribyl and Vanek, 1980; Bruton et al., 1982; Baldis et
al., 1987; Aceñolaza and Aceñolaza, 1992; Tortello and Esteban, 1999;
Tortello et al., 1999). Jujuyaspis is also represented in the
lowermost Tremadocian of Kazakhstan (Balashova, 1961) and Western Australia
(Shergold, 2000).
Classical
lowest Ordovician localities of northwestern Argentina are summarized in this
chapter.
Some
trilobite assemblages have proved to have great value as guides for the base of
the Tremadocian.
The
dominance of olenids enables correlation with deep water successions. Besides,
occurrences of planktic graptolites and conodonts provide further information on
the stratigraphic range and correlation of some sections.
The
Parabolina
(N.) frequens argentina Zone and the position of the Cambrian- Ordovician
boundary
The
Parabolina (Neoparabolina) frequens argentina Biozone
(Harrington and Leanza, 1957) was described on the basis of trilobite faunas
dominated by agnostoids and olenids of cosmopolitan and atlantic aspect.
Although the whole Biozone was originally assigned to the Lower Tremadocian,
Late Cambrian affinities of some species were clearly indicated by Kayser
(1876), Kobayashi (1936, 1937), Harrington and Leanza (1957) and Branisa (1965).
The
lower part of the biozone is characterized by a distinctive latest Cambrian
assemblage mainly composed of Lotagnostus (Lotagnostus), L. (Semagnostus),
L. (Distagnostus), Micragnostus, Strictagnostus,
Pseudorhaptagnostus (Machairagnostus), Gymnagnostus, Leiagnostus,
Beltella, Onychopyge, Plicatolina, Parabolina (Neoparabolina), Parabolinella and
Angelina. These agnostoid-polymeroid faunas are typically recorded in
both the lower part of the Volcancito Formation at río Volcancito and the
Lampazar Formation at sierra de Cajas (Aceñolaza, 1968; Tortello and Aceñolaza,
1993; Esteban, 1999; Tortello and Esteban, 1999), characterizing the Pseudorhaptagnostus
(Machairagnostus) - Gymnagnostus Assemblage Subzone (Tortello,
in press).
Based on a global evaluation of the Lower Paleozoic of South America, Aceñolaza (1983) pointed out that the Cambrian-Ordovician boundary can be defined at the first appearances of Jujuyaspis keideli and Rhabdinopora flabelliformis (Graptolithina), demonstrating that the upper part of the Parabolina (N.) frequens argentina Biozone is assignable to the earliest Tremadocian (see also Salfity
et
al., 1984; Moya et
al., 1994). This scheme coincides with the recent designation of
the base of the Ordovician System by the COBWG (ISOS, ICS, IUGS), which supports
that the FAD of planktic graptolites and Jujuyaspis borealis Kobayashi
constitutes a reasonable proxy for the boundary in North America (Cooper et
al., 2001).
Jujuyaspis
keideli Kobayashi is the most common olenid of the upper part
of the P. (N.) frequens argentina Biozone (Benedetto, 1977). This taxon
is frequently found in association with Parabolinella and Angelina. On
the other hand, the agnostoids are rather scarce in this part of the unit, being
restricted to Trilobagnostus chiushuensis (Kobayashi; Tortello et al.,
1999) and some forms described under open nomenclature (e.g. Gymnagnostus sp.,
Anglagnostus? sp., Micragnostus sp.; Tortello and Aceñolaza,
1999) (Tortello, in press).
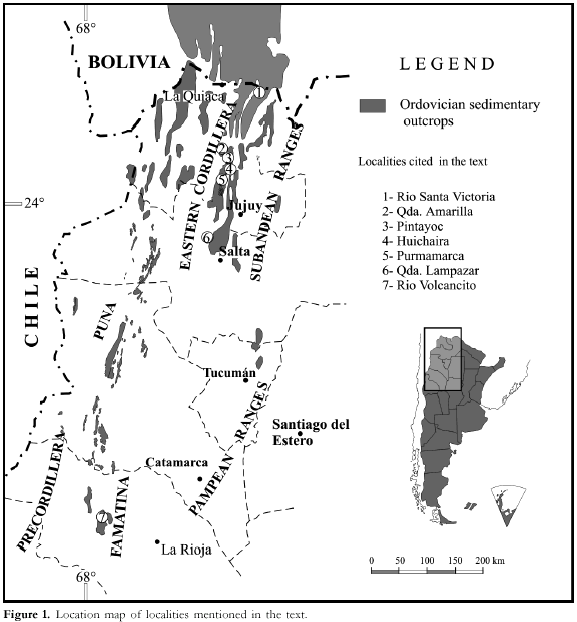
Stratigraphy
Stratigraphic
information on seven classical sections from the lowest Ordovician of the
Cordillera Oriental and Famatina Range (northwestern Argentina; Figure 1) is
briefly summarized below.
The
corresponding correlation chart is shown in Figure 2.
Río
Santa Victoria. The
Santa Rosita Formation is well represented along Rio the Santa Victoria (norhern
Salta Province). This 1.425-m-thick succession is mainly composed of dark grey
to black shales with intercalations of greenish-grey sandstones and shaly
sandstones. The lower 600 m are very fossiliferous, yielding a varied trilobite
fauna assignable to the Parabolina (N.) frequens argentina Biozone. The
Cambrian-Ordovician boundary is recognized by the first appearance of Jujuyaspis
keideli 250 m above the base of the formation, a few meters below the lowest
observed occurrence of Rhabdinopora flabelliformis. On the other hand, Kainella
meridionalis occurs 880 m above the base of the section, in association with
Angelina hyeronimi (Kayser), Leptoplastides marianus (Hoek) and Parabolinella
argentinensis Kobayashi (see Harrington and Leanza, 1957).
Pintayoc.
This 150-m-thick section is located at the quebrada de Sapagua (eastern flank of
Azul Pampa Range, 25 km northwest Humahuaca City). It is mainly composed of
greenish-grayish sandstones and shales assignable to the Chalhualmayoc Formation
(Mesón Group) and the lower part of the Santa Rosita Formation (Santa Victoria
Group). Strata of the upper part of the succession are tectonically disrupted.
The Cambrian-Ordovician boundary is recognized by the occurrence of large sized,
well preserved specimens of Rhabdinopora flabelliformis (Graptolithina).
Associated fossils are represented by trilobites of the Parabolina (N.)
frequens argentina Biozone, brachiopods and echinoderms (see Aceñolaza,
1996a; Aceñolaza, 1998).
Quebrada
Amarilla. The section crops out at the quebrada Amarilla
(northern part of the Cajas Range, Jujuy Province). The 72-m-thick succession is
composed of quartzites, sandstones and marls with intercalations of shales of
the Cardonal Formation. The basal part of the section is assignable to the
latest Cambrian (middle part of the Parabolina (N.) frequens argentina Biozone;
Cordylodus caboti and C. intermedius Biozones). The
Cambrian-Ordovician boundary is recognized by the first appearance of Jujuyaspis
keideli (upper part of the P. (N.) frequens argentina Biozone; Cordylodus
lindstromi Biozone). This species occurs associated with Trilobagnostus
chiushuensis (Kobayashi), Angelina hyeronimi (Kayser) and Pseudokainella
sp. On the other hand, Kainella meridionalis characterizes the
homonymous biozone in the uppermost levels of the section (Cordylodus
angulatus Biozone; late Lower Tremadocian) (see Aceñolaza, 1968; Rao and
Hunicken, 1995; Ortega and Rao, 1995; Rao, 1999; Tortello et al., 1999).
Quebrada
de Huichaira. This
Cambrian-Ordovician section is at Pocoia locality (quebrada de Huichaira, middle
part of the quebrada of Humahuaca). The 232-m-thick succession is composed of
highly fossiliferous green and black shales bearing trilobites, brachiopods and
echinoderms, subordinate sandstones and quartzites. Beds are sliced by
longitudinal faults of a general submeridional trend. The Cambrian-Ordovician
boundary is indicated by the occurrence of Jujuyaspis keideli (Parabolina
(N.) frequens argentina Biozone). The upper part of the section is
assignable to the Kainella meridionalis Biozone (Aceñolaza, 1996b).
Purmamarca. Several fragmentary Cambrian-Ordovician sections display on the western flank of the Sierra Alta (Purmamarca region). This is the type area of the Lower Ordovician index fossil
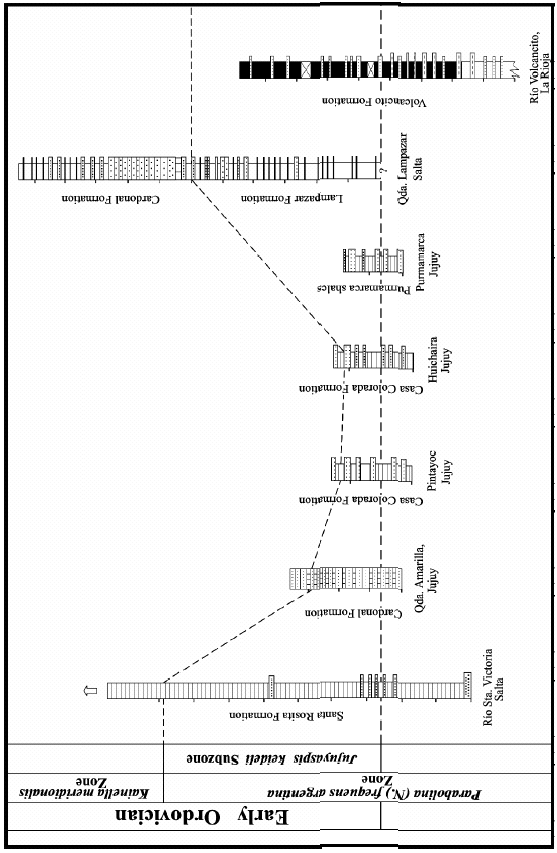
Figure 2. Correlation of Cambro-Ordovician sections in NW Argentina.
Jujuyaspis
keideli (Kobayashi,
1936; Aceñolaza and Aceñolaza, 1992; Miller and Taylor, 1995; Aceñolaza et
al., 2001). Sections
are characterized by anoxic black shales, subordinate sandstones and quartzites.
This
area was strongly affected by tectonics. Shales contain Jujuyaspis keideli and
associated trilobites of the upper part of the Parabolina (N.) frequens
argentina Biozone (Angelina hyeronimi, Parabolinella argentinensis,
Plicatolina sp., Micragnostus sp., Anglagnostus sp.).
“Inarticulate” brachiopods were also described from these strata (Harrington
and Leanza, 1957; Aceñolaza, 1996b; Mángano et al., 1993; Tortello and
Aceñolaza, 1999).
Angosto
de Lampazar. This
succession is located in the Angosto de Lampazar area (eastern flank of the
quebrada del Toro, Salta Province). It represents one of the most classic Lower
Palaeozoic localities in northwestern Argentina. The lower 93 m are shales and
fine-grained sandstones of the Lampazar Formation, which are transitionally
overlain by sandstones, quartzites, marls, shales, and limestones of the
Cardonal Formation (77 m). The Lampazar Fm. bears the trilobites Parabolina
(N.) frequens argentina (Kayser), Leptoplastides marianus (Hoek) and Parabolinella
sp. (P. (N.) frequens argentina Biozone), whereas the middle part of
the Cardonal Fm. is characterized by Leptoplastides marianus (Hoek), Trilobagnostus
chiushuensis (Kobayashi), Asaphellus catamarcensis Kobayashi, Kainella
meridionalis Kobayashi and Pseudokainella conica (Kobayashi) (K.
meridionalis Biozone). In this section, the upper part of the P. frequens
Zone and the basal part of the K. meridionalis Zone correlate with
the Cordylodus angulatus Zone (late Lower Tremadocian) (see Harrington
and Leanza, 1957; Rao and Tortello, 1998; Tortello and Rao, 2000).
Río
Volcancito This section is at the deep Río Volcancito
creek (eastern flank of the Famatina Range, La Rioja Province). The 170 m thick
succession is mainly composed of dark grey and green shales and marls
interbedded with carbonate sandstones of the Volcancito Formation. The lower
part of the unit contains distinctive trilobites (lower part of the Parabolina
(N.) frequens argentina Biozone) and conodonts (Cordylodus proavus, C.
intermedius and C. lindstromi Biozones) of latest Cambrian age. The
Cambrian-Ordovician boundary is recognized by the first appearance of Iapetognathus
aengensis at 85 m from the base of the section, a few meters below the first
occurrences of Jujuyaspis keideli (upper part of the P. (N.) frequens
argentina Biozone) and planktic graptolites (Rhabdinopora flabelliformis Biozone)
(see Harrington and Leanza, 1957; Esteban 1999; Tortello and Esteban, 1997,
1998, 1999; Albanesi et al., 1999, 2000).
The
Jujuyaspis
keideli Kobayashi Subzone
As
noted above, occurrences of Jujuyaspis keideli in the quebrada Amarilla /
Huichaira / Purmamarca and río Volcancito sections provide valuable keys to
characterize the Cambrian-Ordovician boundary in the Cordillera Oriental and the
Famatina Range, respectively. In addition, this species has been described from
many other localities of Argentina and Bolivia (e.g. Tarija, La Caldera, San José,
quebrada de La Cruz; see Harrington and Leanza, 1957; Branisa, 1965; Suárez-Soruco,
1976; Pribyl and Vanek, 1980; Aceñolaza and Aceñolaza, 1992).
Following
the biostratigraphic scheme proposed by Aceñolaza and Baldis (1987), the Jujuyaspis
keideli Subzone is formally proposed here. This new biostratigraphic unit is
based on the stratotype of the río Volcancito section, where the subzone ranges
throughout the upper part of the “Lower Member” of the Volcancito Formation
[upper part of the Parabolina (Neoparabolina) frequens
argentina Biozone] (Esteban, 1999; Tortello and Esteban, 1999). In this
succession, Jujuyaspis keideli is recorded in association with the
graptolites Rhabdinopora flabelliformis cf. socialis (Salter) and Rhabdinopora
flabelliformis cf. norvegica (Bulman) (Esteban, 1999; Tortello and
Esteban, 1999).
The
J. keideli Subzone is correlatable with the graptolite Rhabdinopora
flabelliformis socialis Zone and the conodont Cordylodus lindstromi and
C. angulatus (lower part) Zones (Moya et al., 1994; Ortega y Rao,
1995; Rao, 1999; Albanesi et al., 1999; Tortello and Esteban, 1999).
Representative trilobites from the sections cited above are listed here with
reference to original description:
Angelina
hyeronimi (Kayser,
1876, p. 7, pl. 1, fig. 5).
Anglagnostus?
sp. (Tortello and Aceñolaza, 1999, p. 586, figs. 2.A-B).
Bienvillia
sp. (Aceñolaza
and Aceñolaza, 1992, fig. 2.F).
Jujuyaspis
keideli Kobayashi (1936, p. 90, pl. 16, figs. 5-9).
Leptoplastides
marianus (Hoek, 1912, p. 226, pl. 7, figs. 1-3).
Micragnostus
sp. (Tortello and
Aceñolaza, 1999, p. 587, figs. 2.C-D).
Parabolinella
argentinensis Kobayashi
(1936, p. 88, pl. 15, figs. 1-5).
Pseudokainella
sp. (Tortello et
al., 1999, p.5-566, fig. 4.Q).
Trilobagnostus
chiushuensis (Kobayashi,
1931, p. 173, pl. 22, figs. 1-5).
The
Kainella meridionalis Kobayashi
Zone
Based
on the stratigraphic scheme by Harrington and Leanza (1957), the Parabolina
(N.) frequens argentina Zone is overlain by the Kainella meridionalis Zone.
The latter is widely represented in the Eastern Cordillera (Salta and Jujuy
Provinces), including the classical sections of Santa Victoria, Cajas Range, and
Angosto de Lampazar (Harrington and Leanza, 1957). The unit correlates with the Cordylodus
angulatus and the Rossodus manitouensis Zones (Rao and Hunicken,
1995; Ortega and Rao, 1995; Rao and Tortello, 1998; Rao, 1999; Tortello and Rao,
2000). The most representative trilobites of this zone association include:
Angelina
sp. (Tortello et
al., 1999, p. 5-565, figs. 4.A-F).
Apatokephalus
tibicen Pribyl and Vanek (1980, p. 23, pl. 12, figs. 3-4).
Asaphellus
catamarcencis Kobayashi (1935, p. 65, pl. 11, figs. 11-15).
Brackebuschia
acheila Harrington and Leanza (1957: 226, fig. 123.a-d).
Kainella
meridionalis Kobayashi
(1935, p. 64, pl. 11, figs. 5-9).
Leptoplastides
marianus (Hoek, 1912, p. 226, pl. 7, figs. 1-3).
Pseudokainella
conica (Kobayashi,
1935, p. 65, pl. 11, figs. 16-17).
Trilobagnostus
hoeki (Kobayashi; see Harrington and Leanza, 1957, p. 68,
figs. 11.3-6).
Aknowledgements:
The participation of G.A. was possible with the
finantial support of the Fundación Antorchas.
Authors
aknowledge D. Ruiz Holgado for the line drawings.
References
Aceñolaza,
F.G. 1968. Geología estratigráfica de la región de la Sierra
de Cajas, Dpto. Humahuaca (Provincia de Jujuy). Revista de la Asociación
Geológica Argentina 23 : 207-222.
Aceñolaza,
F.G. 1983. The
Tremadocian beds and the Cambrian-Ordovician boundary problems in Latin America.
En: Papers for the Symposium on the Cambrian-Ordovician and
Ordovician-Silurian Boundaries, Nanjing Institute of Geology and
Palaeontology, Academia Sinica, pp. 88-93.
Aceñolaza,
G.F., 1996a. Presencia de Rhabdinopora (Grpatolithina) en la Quebrada de
Humahuaca, Provincia de Jujuy. Ameghiniana 33 (1): 111-128. Buenos Aires.
Aceñolaza,
G.F. 1996b. [Bioestratigrafía del límite Cámbrico-Ordovícico y Ordovícico
basal en la quebrada de Humahuaca, provincia de Jujuy, República Argentina.
Doctoral. Thesis, Universidad Nacional de Tucumán, 245 pp].
Aceñolaza,
G.F., 1998. Graptolites,
conodonts and trilobites of uppermost Cambrian and Lower Ordovician age from
Jujuy Province, northern Argentina. Temas Geológico-Mineros Instituto Tecnológico Geominero
de España, 23:
145- 147. Madrid.
Aceñolaza,
F.G. and Aceñolaza, G.F. 1992. The genus Jujuyaspis as a world reference fossil
for the Cambrian- Ordovician boundary, p. 115-120. En: B.D. Webby and J.R.
Laurie (eds.), Global Perspectives on Ordovician Geology, Balkema,
Rotterdam.
Aceñolaza,
F.G. and Baldis, B.A.J. 1987. The Ordovician System of South America. IUGS
Publication 22:
1-68.
Aceñolaza,
G.F., Tortello, M.F. and Rábano I., 2001. The eyes of the early Tremadoc Olenid
trilobite Jujuyaspis keideli Kobayashi, 1936. Journal of Paleontology 75
(2): 346-350.
Albanesi,
G.L.; Barnes, C.R. and Hünicken, M.A. 2000. Conodont paleobiogeography of the
Iapetus ocean in the Cambrian-Ordovician boundary interval. XXXI
International Geological Congress,
Río de Janeiro, Artículo en CDroom.
Albanesi,
G.L., Esteban, S.B. and Barnes, C.R. 1999. Conodontes del intervalo del límite Cámbrico-Ordovícico
en la Formación Volcancito, Sistema de Famatina, Argentina. Temas Geológico-Mineros,
Instituto Tecnológico Geominero de España 26: 521-526.
Balashova,
E.A. 1961. Some
Tremadoc trilobites of the Aktyubinsk region. Akademiya Nauk SSSR, Trudy
Geologicheskogo Instituta 18: 102-145.
Baldis,
B.A.J., González, S. and Pérez V. 1984. Trilobites tremadocianos de la Formación
Negritos (Perforación La Hielera), Llanos de Colombia. III Congreso
Latinoamericano de Paleontología (Méjico), Memorias: 28-41.
Benedetto,
J.L. 1977. Algunas
consideraciones acerca de la posición del límite Cambro-Ordovícico en América
del Sur. Geos 23: 3-11.
Branisa,
L. 1965. Los fósiles guías de Bolivia. Boletín Servicio Geológico de
Bolivia (GEOBOL) 6: 1-82.
Bruton,
D.L., Erdtmann, B.D. and Koch, L. 1982. The Naersnes section, Oslo Region,
Norway: a candidate for the Cambrian-Ordovician boundary stratotype at the base
of the Tremadoc Series. En: M.G. Basset and W.T. Dean (eds.), The
Cambrian-Ordovician boundary: sections, fossils distributions, and correlations,
National Museum of Wales, Geological Series 3, Cardiff, pp. 61-69.
Chen,
J., Zhou, Z., Zou, X., Lin, Y., Yang, X., Wang, Q., Qi, J., Wang, Q. and Lu, X.
1980. Ordovician sediments and faunas in the Taihang Mountains, North China. Memoirs
of Nanjing Institute of Geology and Palaeontology, Academia Sinica 16:
111-148.
Cooper,
R.A., Nowlan, G.S. and Williams, S.H. 2001. Global Stratotype Section and Point
for base of the Ordovician System. Episodes 24(1 : 19-28.
Dean,
W.T. 1989. Trilobites from the Survey Peak, Outram and Skoki Formations (Upper
Cambrian-Lower Ordovician) at Wilcox Pass, Jasper National Park, Alberta. Geological
Survey of Canada Bulletin 389: 1-141.
Esteban,
S.B. 1999. [Estratigrafía, geología sedimentaria y paleontología del Ordovícico
basal del Sistema de Famatina. Unpublished Ph.D. thesis, Facultad de
Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán,
Argentina].
Fortey,
R.A., Landing, E. and Skevington, D. 1982. Cambrian-Ordovician boundary sections
in the Cow-Head Group, Western Newfoundland. En: M.G. Basset and W.T. Dean
(eds.). The Cambrian-Ordovician boundary: sections, fossils distributions,
and correlations, National Museum of Wales, Geological Series, 3, Cardiff,
pp. 95-129.
Harrington,
H.J. and Leanza, A.F. 1957. Ordovician trilobites of Argentina. Department of
Geology, University of Kansas Special Publication 1: 1-276.
Henningsmoen,
G. 1957. The trilobite family Olenidae, with description of Norwegian material
and remarks on the Olenid and Tremadocian Series. Skrifter Utgitt av det
Norske Videnskaps-Akademi i Oslo I Matematisk- Naturvidenskapelig Klasse 1957(1),
pp. 303, 31 pl.
Kayser,
E. 1876. Über primordiale und untersilurische Fossilien aus der Argentinischen
Republik. Palaeontographica, Supplementum 3, Lieferung 2, Theil
II.
Kim,
D.H. and Choi, D.K. 2000. Jujuyaspis and associated trilobites from the
Mungok Formation (Lower Ordovician), Yongwol, Korea. Journal of Paleontology 74:
1031-1042.
Kobayashi,
T. 1931. Studies on the Stratigraphy and Palaeontology of the Cambro-Ordovician
Formation of Hua- Lien-Chai and Niu-Hsin-Tai, south Manchuria. Japanese
Journal of Geology and Geography 8 (3): 131-189, pls. 16-22.
Kobayahi,
T. 1935. On the Kainella fauna of the Basal Ordovician age found in
Argentina. Japanese Journal of Geology and Geography 12 (3-4): 59-67.
Kobayashi,
T. 1936. On the Parabolinella Fauna from Province Jujuy, Argentina with a
note on the Olenidae. Japanese Journal of Geology and Geography 13 (1-2):
85-102, pls. 15-16.
Kobayashi,
T. 1937. The Cambro-Ordovician shelly faunas of South America. Journal of the
Faculty of Science, Imperial University of Tokyo, Section 2, 4 (4): 369-522,
8 pl.
Loch,
J.D., Stitt, J.H. and Miller, J.F. 1999. Trilobite biostratigraphy through the
Cambrian-Ordovician boundary interval at Lawson Cove, Ibex, western Utah,
U.S.A.. Acta
Universitatis Carolinae Geologica 43: 13-16.
Miller,
J.F. and Stitt, J.H. 1995. Stratigraphic position and significance of Jujuyaspis
and Iapetognathus in the Wilberns Formation, Texas. Ordovician
Odyssey: Short Papers for the seventh International Symposium on the Ordovician
System (Las Vegas), pp. 105-108.
Miller,
J.F. and Taylor, M.E. 1995. Biostratigraphic significance of Iapetognathus (Conodonta)
and Jujuyaspis (Trilobita) in the House Limestone, Ibex area, Utah. Ordovician
Odyssey: Short Papers for the seventh International Symposium on the Ordovician
System (Las Vegas), pp. 109-112.
Moya,
M.C., Malanca, S., Monteros, J.A. and Cuerda, A.J. 1994. Bioestratigrafía del
Ordovícico Inferior en la Cordillera Oriental Argentina basada en graptolitos. Revista
Española de Paleontología 9: 91-104.
Norford,
B.S. 1969. The Early Canadian (Tremadocian) trilobites Clelandia and Jujuyaspis
from the Southern Rocky Mountains of Canada. Geological Survey of Canada,
Contributions to Canadian Paleontology, Bulletin 182: 1-15, 56-59.
Ortega,
G. and Rao, R.I. 1995. Nuevos
hallazgos de graptolitos (Ordovícico Inferior) en la sierra de Cajas,
Departamento Humahuaca, Provincia de Jujuy, Argentina. Boletín de la
Academia Nacional de Ciencias 60: 293-316.
Pribyl,
A. and Vanek, J. 1980. Ordovician
trilobites of Bolivia. Rozpravy Ceskoslovenské Akademie Ved 90(2), 90
pp.
Rao,
R.I. 1999. Los conodontes Cambro-Ordovícicos de la sierra de Cajas y del
Espinazo del Diablo, Cordillera Oriental, República Argentina. Revista Española
de Micropaleontología 31: 23-51.
Rao,
R.I. and Hünicken, M.A. 1995. Conodont biostratigraphy of the
Cambrian-Ordovician boundary in northwestern Argentina. Ordovician Odyssey:
Short Papers for the seventh International Symposium on the Ordovician System (Las
Vegas), pp. 125-128.
Rao,
R.I. and Tortello, M.F. 1998. Tremadoc conodonts and trilobites from the
Cardonal Formation, Incamayo Creek, Salta Province, northwestern Argentina. Palaeontologia
Polonica 58:
31-45.
Salfity,
J.A., Malanca, S., Moya, M.C., Monaldi, C.R. and Brandán, E.M. 1984. El límite
Cámbrico-Ordovícico en el norte de la Argentina. IX Congreso Geológico
Argentino, Actas 1: 568-575.
Shergold,
J.H. 2000. The Early Ordovician trilobite genus Jujuyaspis in Australia.
En: G.F. Aceñolaza, and S. Peralta (Eds.), Cambrian from the southern edge,
INSUGEO, Miscelánea 6: 128-130.
Steinmann,
G. and Hoek, H. 1912. Das
Silur und Cambriun des Hochlandes von Bolivia und ihre Fauna. Neues
Jahrbuch für Mineralogie, Geologie und Paläontologie 34,
176-252.
Stitt,
J.H. and Miller, J.F. 1987. Jujuyaspis borealis and associated trilobites
and conodonts from the Lower Ordovician of Texas and Utah. Journal of
Paleontology 61: 112-121.
Suárez
Soruco, R. 1976. El Sistema Ordovícico en Bolivia. Revista Técnica de YPFB 5
(2): 1-223.
Taylor,
J.F. and Repetski, J.E. 1995. High-resolution trilobite and conodont
biostratigraphy across the Cambrian- Ordovician boundary in south-central New
Mexico. Ordovician Odyssey: Short Papers for the seventh International
Symposium on the Ordovician System (Las Vegas), pp. 133-136.
Tortello,
M.F. (in press). Biostratigraphic significance of the uppermost Cambrian-lowest
Ordovician agnostoid trilobites from northwestern Argentina. Acta
Geológica Hispánica.
Tortello,
M.F. and Aceñolaza, G.F. 1993. Trilobites agnóstidos del límite Cámbrico-Ordovícico
de la Formación Lampazar, Sierra de Cajas, Provincia de Jujuy, Argentina. Revista
Española de Paleontología 8 (2): 177-184.
Tortello,
M.F. and Aceñolaza, G.F. 1999. Trilobites agnóstidos del Ordovícico basal en
la localidad de Purmamarca, Provincia de Jujuy, Argentina. Temas Geológico-Mineros,
Instituto Tecnológico Geominero de España 26: 585-588.
Tortello,
M.F. and Esteban, S.B. 1997. Significado bioestratigráfico de una asociación
de trilobites del tramo basal de la Formación Volcancito (Sistema de Famatina,
La Rioja, Argentina). Ameghiniana 34 (3): 265-270.
Tortello,
M.F. and Esteban, S.B. 1998. La transición Cámbrico-Ordovícica en la Formación
Volcancito (Sistema de Famatina, La Rioja, Argentina). VII Congreso Argentino
de Paleontología y Bioestratigrafía, Resúmenes: 93.
Tortello,
M.F. and Esteban, S.B. 1999. La transición Cámbrico-Ordovícico en la Formación
Volcancito (sierra de Famatina, La Rioja, Argentina). Ameghiniana 36:
371-387.
Tortello,
M.F., Rábano, I., Rao, R.I. and Aceñolaza, F.G. 1999. Los trilobites de la
transición Cámbrico-Ordovícico en la quebrada Amarilla (sierra de Cajas,
Jujuy, Argentina). Boletín Geológico y Minero 110: 555-572.
Tortello,
M.F. and Rao, R.I. 2000. Trilobites
y conodontes del Ordovícico Temprano del Angosto de Lampazar (provincia de
Salta, Argentina). Boletín Geológico y Minero 111: 61-84.
Westrop,
S.R. 1986. Trilobites of the Upper Cambrian Sunwaptan Stage, southern Canadian
Rocky Mountains, Alberta. Palaeontographica Canadiana 3: 1-179.
Winston,
D. and Nicholls, H. 1967. Late Cambrian and Early Ordovician Faunas from the
Wilberns Formation of Central Texas. Journal of Paleontology 41: 66-96.
Zhou,
Z.Y. and Zhang, J.L. 1984. Uppermost Cambrian and lowest Ordovician trilobites
of Northeast China. En: Nanjing Institute of Geology and Palaeontology,
Stratigraphy and Palaeontology of Systemic Boundaries in China, Cambrian-
Ordovician Boundary (2), Anhui Science and Technology Publishing
House, Hefei, pp. 63-163.
Recibido:
12 de Octubre de 2002
Aceptado: 4 de Diciembre de 2002
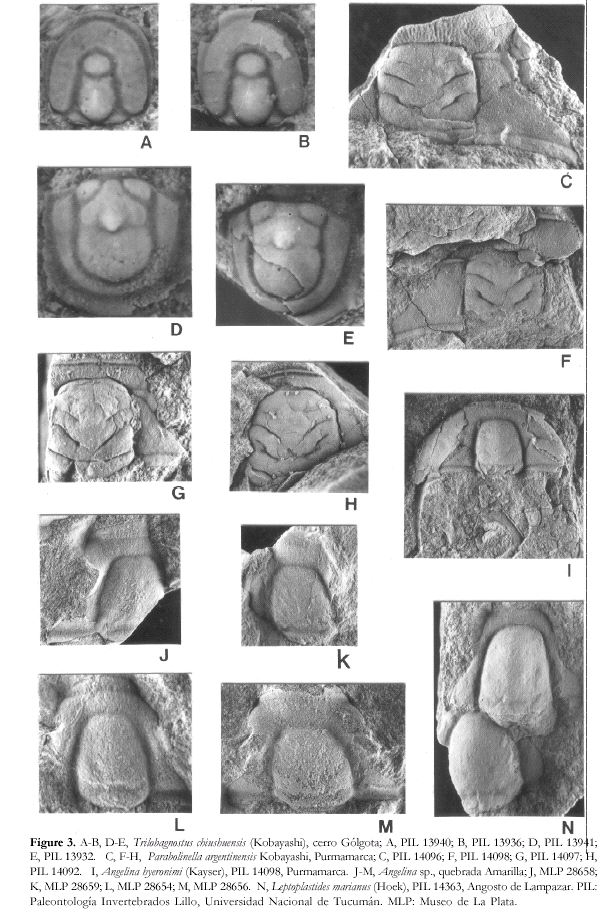
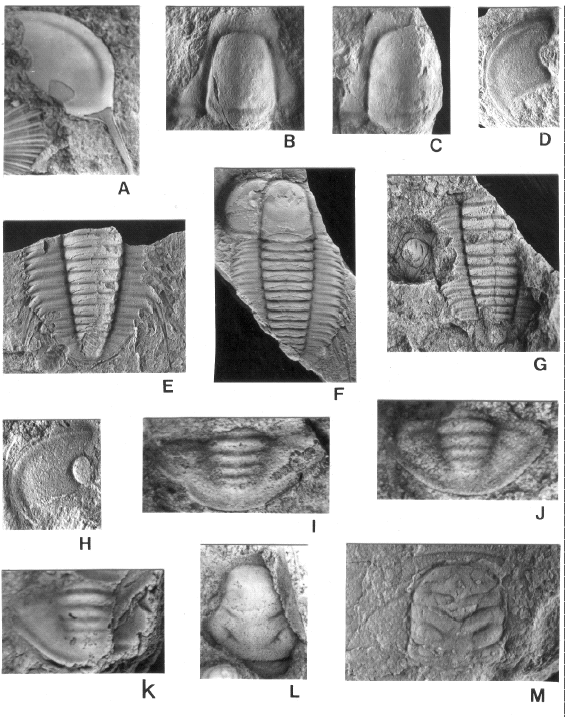
Figure 4.
A-C, Leptoplastides marianus (Hoek); A, PIL 13936, cerro Gólgota; B, PIL 14363, Angosto de Lampazar; C, PIL 14361, Angosto de Lampazar. D-K, Jujuyaspis keideli Kobayashi; D, MLP 28603, quebrada Amarilla; E, MLP 30292, Purmamarca; F, MLP 30269, Purmamarca; G, PIL 14092, Purmamarca; H, MLP 28603, quebrada Amarilla; I, MLP 28612, quebrada Amarilla; J, MLP28619, quebrada Amarilla; K, MLP 28622, quebrada Amarilla. L, Apatokephalus tibicen Pribyl and Vanek, MLP 28662, quebrada Amarilla. M, Plicatolina sp., PIL 14099, Purmamarca. PIL: Paleontología Invertebrados Lillo, Universidad Nacional de Tucumán. MLP: Museo de La Plata.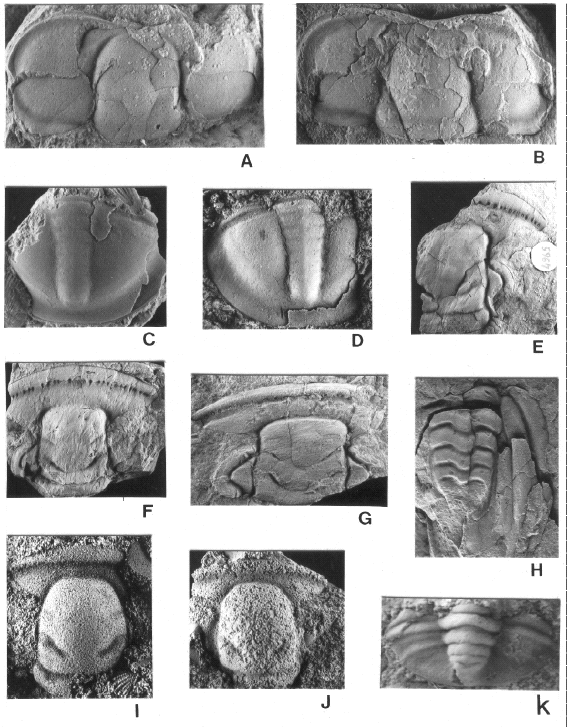
Figure 5
: A-B, Jujuyaspis keideli Kobayashi, Purmamarca; A, MLP 30289; B, MLP 30351. C-D, Asaphellus catamarcencis Kobayashi, Angosto de Lampazar; C, MLP 28768; D, MLP 28769. E-H, Kainella meridionalis Kobayashi, Iruya; E, MLP 4965; F, MLP 4957; G, MLP 4929; H, MLP 4954. I-J, Pseudokainella conica (Kobayashi), Angosto de Lampazar; I, MLP 28849; J, MLP 28851. K, Brackebuschia acheila Harrington and Leanza, PIL 14087, cerro Gólgota. PIL: Paleontología Invertebrados Lillo, Universidad Nacional de Tucumán. MLP: Museo de La Plata.