Sponge Fauna and Spicule Assemblages from the
Ordovician of Argentina: A Review
Matilde S. BERESI1 and Susana B. ESTEBAN2
1 CRICYT-IANIGLA. Avda. A.Ruiz Leal s/n, 5500 Mendoza, E-mail: mberesi@lab.cricyt.edu.ar
2 INSUGEO - Facultad de Cs. Naturales e I.M.L, Miguel Lillo 205, 4000 Tucumán, E-mail: insugeo@unt.edu.ar
Abstract.
SPONGE
FAUNA AND SPICULE ASSEMBLAGES FROM THE ORDOVICIAN
OF ARGENTINA:
A REVIEW.
Ordovician
sponges and sponge spicules assemblages were described from many localities of
Precordillera; Famatina and Puna, north of Argentina. The most diverse and
well-preserved faunas are from the San Juan Formation (Arenig-Lower Llanvirn,
Precordillera terrane, western Argentina). Precordilleran sponge faunas are
dominated by orchoclad lithistid demosponge genera, whereas hexactinellids are
represented by loose spicules and root tufts, and calcareous heteractinid
sponges are known from isolated octactine spicules and a single genus. In
addition, spicules assemblages were reported from diverse localities of the
Precordillera, a hexactinellid sponge genus was described from the Tremadocian
of the Puna area, and hexactinellid spicule assemblages were reported from the
lower Tremadocian black shales of the Famatina System. A discussion on the
paleoecological and paleoambiental implications of these faunas is given.
Resumen.
LA
FAUNA DE ESPONJAS Y ASOCIACIONES DE ESPÍCULAS DEL ORDOVÍCICO
DE ARGENTINA:
UNA REVISIÓN.
Las esponjas y las
asociaciones de espículas mencionadas en este trabajo fueron coleccionadas en
diversas localidades de Precordillera, Sistema de Famatina y Puna, en el norte
de Argentina. La fauna de esponjas más diversa y mejor preservada del Ordovícico
de Argentina proviene de la Formación San Juan (Arenigiano-Llanvirniano
inferior, terreno de Precordillera, oeste de Argentina). La fauna de esponjas
precordilleranas está dominada por géneros de demospongeas lithístidas del
Suborden Orchocladina, aunque hexactinéllidas son conocidas mediante espículas
sueltas y penachos de raíces. Esponjas heteractínidas calcáreas están
representadas por un sólo género y por espículas octactinas aisladas.
Asociaciones de espículas fueron registradas de diversas localidades de
Precordillera. Un género de esponja hexactinéllida fue descripto de las
sedimentitas volcaniclásticas tremadocianas de la Puna, en tanto que
asociaciones de espículas hexactinéllidas fueron colectadas en las lutitas
negras del Tremadociano inferior del Sistema de Famatina. Se presenta un resumen
de las consideraciones paleoecológicas y paleoambientales.
Key words: Ordovician.
Sponges. Puna.
Famatina. Precordillera.
Palabras
clave: Ordovícico.
Esponjas. Puna. Famatina. Precordillera.
Introduction
Sponges are the most primitively organized
multicellular animals. Specialized cells of most sponges secrete a mineral
skeleton (spicules) which provides a firm basis for classification.
The Ordovician sponges from Argentina are mostly
represented in the Precordillera terrane (western Argentina). Well-preserved and
diverse faunas occur in the carbonate platform of the San Juan Formation
(Arenig-Lower Llanvirn) in the San Juan Province, constituting the most
extensive record of Ordovician sponges from South America.
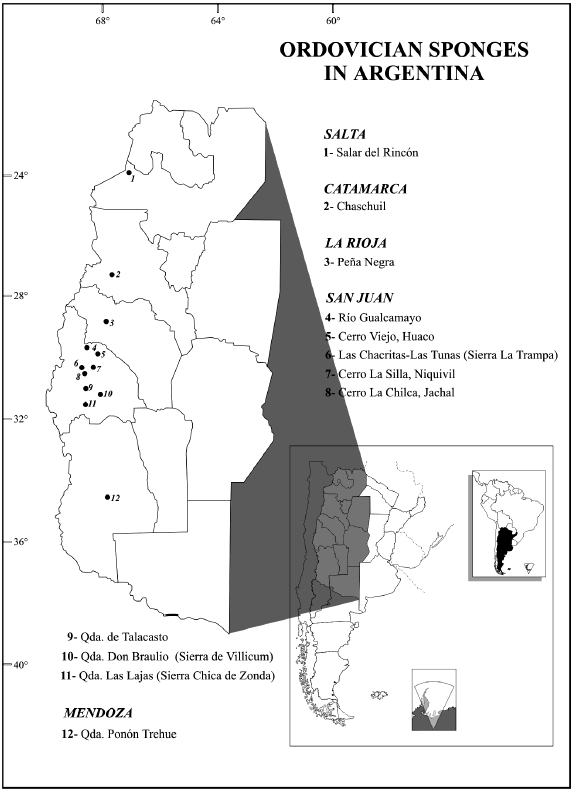
In addition, fragmentary remains of Ordovician spicules have been reported in the Famatina System and the Puna area (northwestern Argentina): isolated hexactinellid spicules were mentioned from the Chaschuil area (Catamarca Province) (Aceñolaza and Toselli, 1977), fragments of hexactinellid reticulated nets were described from the shales of the Volcancito Formation (Esteban and Rigby, 1998), and a single hexactinellid sponge was found in the Puna area (Carrera, 1998).
Kayser (1925) found sponges at quebrada de Talacasto,
San Juan Precordillera, but they were only mentioned as undetermined
“spongiae”. The first sponge spicules assemblage from the lower Ordovician
of the San Juan Precordillera was fully described by Gnolli and Serpagli (1980).
Micritic beds with Archaeoscyphia in growth position from the upper part
of the San Juan Formation at quebrada de Talacasto were first interpreted as
“buildups” by Beresi (1986). Sponges in situ also occur at Cerro San Roque (Jáchal) and
quebrada Las Lajas (sierra Chica de Zonda). These
Lower- Middle Ordovician buildups were later on described as reef-mound
structures by Keller and Bordonaro (1993) and included in the Archaeoscyphia-Calathium
association (Cañas and Carrera, 1993) and the presence of
stromatoporoid-like organism (zondarella Keller and Flügel, 1996) The
Precordilleran sponge faunas have been intensely studied in the last decades
(Beresi, 1985, 1986, 1990; Beresi and Rigby, 1993; Carrera 1985, 1994a, 1994b,
1996). The distribution pattern of these sponges has been recently analyzed by
Carrera and Rigby (1999) and Carrera (2000, 2001). In addition, Mehl and Lehnert
(1997) reported Cambro-Ordovician sponge spicules assemblages from diverse
localities of the Precordillera, and Beresi and Heredia (2000) have described
sponge spicules assemblages from Lower and Middle Ordovician strata of the
Sierra Pintada, southern Mendoza Province.
The purpose of this paper is to summarize all published
data on the occurrence of Ordovician sponges from Argentina. It should be
pointed out that the descriptions and interpretations offered simply transcribe
the authors’ opinions in the original papers.
Geographic distribution of taxa
The best-know Ordovician sponges come from
threegeographic basins: Puna, Famatina System and Precordillera, from levels
that range from Tremadocian to Llanvirn.
I. Puna
A single specimen of a complete hexactinellid sponge is
the first Ordovician sponge discovered from the Puna region. It has been
collected from volcaniclastic rocks of the Las Vicuñas Formation (Tremadocian)
in quebrada Lari (southwest of the Salar del Rincon area, Salta Province)
(Locality 1 of Figure 1). This specimen was assigned to Larispongia
magdalenae, constituting the first representative of the Family
Pelicaspongiidae in western Gondwana (Carrera, 1998). It is characterized by an
irregular, thick-walled skeleton and large, smooth hexactines on the dermal and
gastral surfaces, which are not arranged into a quadrate pattern. The associated
fauna is represented by trilobites and brachiopods of Tremadocian age.
II.
Famatina System
1.
Catamarca Province
The only discovery of Ordovician sponges in this
province corresponds to isolated hexactinellid spicules described by Aceñolaza
and Toselli (1977) from the Chaschuil region (Locality 2 of Figure 1). The
material appears dispersed in carbonate concretions of the Suri Formation
(Arenig).
2. La Rioja Province
The sponge material came from black siliceous
graptolitic shales of the upper part of the Volcancito Formation (= upper member
of Turner, 1964) at Peña Negra locality (río Achavil, Sierra de Famatina)
(Locality 3 of Figure 1). The sponges are associated with planktic graptolites (Rhabdinopora
and Anisograptus) and remains of dendroid graptolites. Based on the
graptolitic fauna, these levels were assigned to the Lower Tremadocian (Esteban
and Gutierrez-Marco, 1997). The sponges from the Volcancito Formation were
studied by Esteban and Rigby (1998), who recognized fragments of reticulated
skeletal nets of the hexactinellid sponge Protospongia. Because of
incomplete preservation, specific identification was not possible.
III. Precordillera of San Juan Province San
Juan Formation: Upper Tremadocian - Early Llanvirn
1. Villicum Range
The limestone section of quebrada Don Braulio is
exposed on the eastern flank of the Sierra de Villicum (Locality 10 of Figure 1)
about 40 km northeast of San Juan City. The San Juan Formation is 250 m thick at
the Sierra de Villicum. In quebrada Don Braulio, the sponge beds are in the Amorphognathus
variabilis-Eoplacognathus suecicus Zone (Sarmiento, 1985), indicating that
they are Lower Llanvirn in age. These beds are in the upper part of the San Juan
Formation at quebrada Don Braulio. The shaly limestone member is 10 m thick. It
consists of fossiliferous, fine-grained limestones and calcipelitic beds.
The sponge fauna from the Sierra de Villicum locality
includes Hudsonospongia cyclostoma Raymond and Okulitch 1940, Calycocoelia
perforata, Aulocopium sanjuanensis, and Psarodictyum magna. These
orchocladine demosponges occur with hexactine spicules, extensive hexactinellid
root tufts, and an additional single isolated octactine spicule that documents
the presence of the Heteractinida (Beresi and Rigby, 1993).
A shelly fauna composed by orthocean brachiopods,
trilobites, and crinoids is common in the fine limestones. The lithistid sponges
are associated with the other fossils, but root tufts do not occur associated
with other sponges at the quebrada Don Braulio locality.
2.
Huaco area
a.
Buenaventura Luna section
The section studied crops out on the eastern flank of
the core of the Huaco anticline, along the sulphurous quebrada Agua Hedionda.
There, the San Juan Formation is about 170 m thick (Locality 5 of Figure 1).
Sponges occur principally in the shaly limestone member
at the top of the limestone sequence, in association with conodont faunas of
early Upper Arenig age (Baltoniodus navis Zone, Lemos, 1981).
Protachilleum kayseri Zittel
1877 and Archeoscyphia nana Beresi and Rigby 1993, are the only sponges
known at present from the Buenaventura Luna section, in the Huaco locality. They
occur with monactine and hexactine spicules in the dark limestone member,
associated with articulate brachiopods, trilobites, bryozoans, crinoids and
microalgae in marly limestones. Sponges represent only 10 percent of the fossils
in this locality.
b.
Cerro Viejo section
In the Cerro Viejo section (Locality 5 of Figure 1)
nodular limestones and marls of the San Juan Formation contain a diverse sponge
fauna. The conodont fauna is assignable to the Eoplacognathus suecicus Zone
of early Llanvirn age (Hünicken and Ortega, 1987). The sponges are partly or
wholly embedded in irregular lenses surrounded by dark grey micrite from the
mudstone and wackestone layers of the San Juan Formation.
Lithistid demosponges Archaeoscyphia, Calycocoelia and
Hudsonspongia (Carrera 1985), Archaeoscyphia tenuis and root tufts
were recognized (Carrera, 1994a). A new megamorinid genus Rugospongia
viejoensis of the Family Saccospongidae was described from this section
(Carrera, 1996b).
3.
Cerro La Silla, Niquivil
The carbonate sequence consists of skeletal wackestones
alternating with biolithoclastic grainstones (Locality 7 of Figure 1). The
sponge fauna is associated with conodonts assignable to the Oepikodus evae Zone
(Hünicken, 1985) of early Arenig age.
In this section Eospongia and Nexospongia
sillaensis, which is included in the Family Nexospongia, were described by
Carrera (1996b). According to this author, Nexospongia sillaensis is
composed of irregularly distributed heloclones and monaxons as principal
spicules.
4. Cerro La Chilca
In the Cerro La Chilca section (south Jáchal)
(Locality 8 of Figure 1), sponges were collected from skeletal wackestones and
packstones alternating with thin layers of bioclastics grainstones and
rudstones. In this limestones section two new genera and species were described:
the calcareous heteractinid Chilcaia bimuralis (Carrera, 1994a) and a
lithistid species Incrassospongia ramis (Carrera, 1996a) from the upper
levels of the San Juan Formation (early Llanvirn; Eoplacognathus
variabilis-E. suecicus Zones).
Other common sponge genera recognized are Archaeoscyphia,
Patellispongia cf. P. occulata Bassler 1927, as well as root tufts in
life position (Carrera, 1994b).
5. Quebrada de Talacasto
Quebrada de Talacasto is situated in San Juan province,
about 89 km northwest of San Juan City in the Precordillera Central (Locality 9
of Figure 1). The San Juan Formation section is 350 m thick.
Sponge faunas occur in the calcareous shaly limestone
member, in the lower part of the formation, and in the fossiliferous member
(mudstones and skeletal wackestones) at the top of the calcareous sequences. The
lowest conodont faunas correspond to the Oepikodus evae Zone (Hünicken,
1985).
The Pygodus serra Zone occurs in the upper
section. The age of the San Juan Formation at Quebrada de Talacasto ranges from
late Lower Arenig to the Lower Llanvirn.
Quebrada de Talacasto is the most productive Ordovician sponge locality yet known from Argentina. The sponge fauna is one of the most characteristic features in the Ordovician limestones of the San Juan Formation.
The
species from the Talacasto section include: Archaeoscyphia annulata Cullison
1944 (Beresi, 1985), Archaeoscyphia sp., Patellispongia sp.,
Calycocoelia sp. and Nevadocoelia sp. (Beresi, 1986). The
species Archaeoscyphia minganensis (Billings 1859) and Rhopalocoelia
clarkii Raymond and Okulitch 1940 were reported by Beresi and Rigby (1993).
The
new genus and species Talacastonia chela are described from this
locality. The new species from the Talacasto section include: Anthaspidella
inornata, Anthaspidella annulata, Anthaspidella alveola, Archaeoscyphia nana,
Aulocopium sanjuanensis, Hudsonospongia talacastensis, Patellispongia robusta, and
Rhopalocoelia rama (Beresi and Rigby, 1993).
Root
tufts and isolated hexactine spicules are the only known representatives of the
Hexactinellida and a single loose octactine spicule is the only evidence of the
Heteractinida at this locality.
Patellispongia
argentina, Patellispongia cf P. occulata Bassler
1927, Archaeoscyphia tenuis and scarce root tufts were also described
from this section (Carrera, 1994a).
6.
Las Chacritas and Las Tunas sections
Data
on the occurrence of sponge faunas from the carbonate and siliciclastic
sequences overlaying the San Juan Formation are scarce. Sponges have only been
described from two sections of the central Precordillera of San Juan.
In
the Las Chacritas river and the Las Tunas sections (Sierra La Trampa) (Locality
6 of Figure 1), a late Arenig-early Llanvirn carbonate unite is recognised,
bearing conodonts of the Lenodus variabilis Zone. Such unite named Las
Chacritas Formation (Peralta et al., 1999) 70 m in thickness, is composed
mainly of carbonate deposits, overlaying in stratigraphic discontinuity
(paraconformity) the limestones of the San Juan Formation, which is Arenig in
age.
The
collected material comes from the upper part of the carbonate sequence in Las
Chacritas and Las Tunas sections (Carrera, 1997). The sponge association is
mainly composed of two species of the genus Archaeoscyphia: A. pulchra Bassler
and A. minganensis Billings reaching up to 76 % of the sponge biovolume.
Other less represented taxa are: Rhopalocoelia clarkii Raymond and
Okulitch, Hudsonospongia sp., Calycocoelia sp., “root-tufts”
and domical to discoidal sponges.
Sponge
spicule assemblages
The
oldest spicules assemblage from the Lower Ordovician limestones (O.
intermedius Zone) of the San Juan Formation was documented by Gnoli and
Serpagli (1980) in the Pachaco section, San Juan Precordillera. The sponge
spicules were recovered from acetic acid-etched residues for the recovery of
conodonts. The five basic types of spicules found are: undistorted hexactines,
desmas (dencroclones and rhizoclones), octactines and monaxones.
Well
preserved silicified sponge spicule assemblages were described from residues of
conodont samples by Mehl and Lehnert (1997). The Ordovician spicules were
recovered from San Juan, Gualcamayo and Las Aguaditas Formations. The spicule
assemblage includes Polyactinellidae, Heteractinellidae (Calcarea) and
hexactinellid and demospongid spicules.
In
the San Juan Formation the assemblages are related to reef-mound horizons and
biostromes with sponges, stromatoporoids, receptaculitids and some autrotrophic
organisms. The genera Dodecaactinella oncera, Sardospongia cynodonta,
Praephobetractinia sp. and Eiffelia sp. were reported from Arenig
strata of the San Juan Formation (Mehl and Lehnert, 1997). From the Gualcamayo
and the upper part of the Las Aguaditas Formations, only hexactine spicules were
documented.
Isolated sponge spicules were recovered from dilute acetic-acid residues of conodonts of clear

Plate
I
1.
Psarodictyum magna Beresi and Rigby, 1994, holotype, nature of the coarse
skeletal net of trabs interrupted by limited canals of the palmate or frond-like
species, San Juan Formation, Villicúm Range, Ianigla PI VI- 44.
2.
Aulocopium sanjuanensis Beresi and Rigby, 1994, oblique side view shows
the general obconical form of the species, Ianigla PI VI-13.
3.
Patellispongia robusta Beresi and Rigby, 1994, side view of obconical,
moderately thin-walled, paratype shows the nature of the skeletal net, San Juan
Formation, Talacasto Gulch section, Ianigla PI T-1/15.
4.
Calycocoelia perforata Beresi and Rigby, 1994, paratype shows the smooth
dermal surface of the subcylindrical species, San Juan Formation, Villicum
Range, Ianigla PI VI-2.
5.
Archaeoscyphia minganensis (Billings, 1859). moderately robust specimen
showing irregularity in annulations, San Juan Formation at Talacasto Gulch,
Ianigla PI T-47.
6.
Anthaspidella annulata Beresi and Rigby, 1994, holotype dermal surface
showing annulate nature of the sponge, Talacasto Gulch section. Ianigla PI T-49.
7.
Hudsonospongia cyclostoma Raymond and Okulitch, 1940, from the Villicum
Range section, showing the general growth form of the species and the moderately
coarse trab-based anthaspidellid skeleton, Ianigla PI VI-2.
gray
limestone and black argillaceous limestone of the Ponón Trehue section, Sierra
Pintada (Locality 12 of Figure 1), southern Mendoza Province, Argentina.
Poriferan taxa include heteractinida spicules as well as hexactinellid
hexactines and non-lithistid demospongiid triaene and oxeas with some doubt.
The
heteractinid spicules were collected from Arenig allochthonous megaconglomerates
of the Oepikodus evae Zone, on the Quebrada Ponón Trehue. Hexactinellid
hexactines and non-lithistid demospongiid triaene and oxeas were described from
Llanvirn autochthonous limestone and carbonate sandstone with the Pygodus
serra Zone and the P. anserinus Zone (Heredia, 1996; 2001) in
the La Tortuga section (southern Quebrada Ponón Trehue).
The
Precordilleran sponge fauna: Paleogeographic and paleoecologic considerations
The
Ordovician sponge faunas from the Precordillera are most similar to assemblages
from North America, particularly to those from the northern Appalachian region
(Beresi and Rigby, 1993; Carrera and Rigby, 1999) and the Great Basin area.
The
lithistid Ordovician sponges are well known from North America, Australia,
Europe, Asia and South America. Generalized genera, such as Archaeoscyphia,
Patellispongia, Aulocopium, Hudsonospongia, and Calycocoelia occur
widely in the Middle Ordovician. Most other genera, however, show some
provincialism. In the Precordillera, endemic genera such as Protachilleum,
Talacastonia, Rugospongia and Chilcaia were reported from the San
Juan Formation. In this way, based on the distribution of endemic and restricted
genera in the Middle Ordovician, the sponge associations from the Precordillera
are included in the “eastern North American Laurentian faunas” (Carrera and
Rigby, 1999).
Some
kinds of sponges (such as Archaeoscyphia) are common reef formers. The
characteristic association Archaeoscyphia, Calathium and
stromatoporoid-like organisms occurs in a broad belt along the southern and
western margins in eastern North America, the Argentina Precordillera and China.
The presence of these reef-mound structures, was interpreted as a characteristic
world wide episode in the early Ordovician apparently restricted to tropical or
subtropical areas without any other paleogeographic significance (Carrera and
Rigby, 1999).
The
evolution of the Ordovician sponge fauna from the Precordillera involves
notorious changes from algal-sponge (reef ecosystems) in the early Arenig to
stromatoporid associations in the Middle Arenig to anthaspidellid demosponge
dominated associations in the Upper Arenig to Lower Llanvirn.
From
the Llanvirn up to the Upper Ordovician, effect of diverse abiotic factors such
as volcanigenic activity, sea level fluctuations and finally the global climatic
cooling could have been contributed to the decrease of the sponges.
Distribution
patterns of the sponges of the Argentine Precordillera were first analyzed by
Beresi and Rigby (1993) and recently by Carrera and Rigby (1999) and Carrera,
2000. These patterns suggest that distributions of sponges were strongly
influenced by depth, current intensity on the carbonate platform and by
continental convergence and latitudinal climatic gradients.
The diverse San Juan sponge faunas of siliceous lithistid demosponges and hexactinellids thrived in an open subtidal environment, but were not successful in shallower, possibly more turbid waters. They are associated with a well preserved biota of brachiopods, bryozoans, nautiloids, gastropods, ostracods, trilobites, crinoids, and algae. The sediments and diversity and abundance of other elements of the biota, in addition to the sponges, suggest good water circulation in an open shelf, a mainly subtidal environment of carbonate deposition, and waters of normal salinity in

Plate
II
1.
Protospongia sp. B
Esteban and Rigby, 1998, specimen PIL 14.193 from Volcancito Formation, Famatina
region, photomicrograph showing spicules and quadrules in the fragment of a
reticulate net.
2.
Protospongia sp. A Esteban and Rigby, 1998, specimen PIL 14.192 from
Volcancito Formation, Famatina region, photomicrograph of skeletal structure
showing relief features on hexactines in the well-ordered skeleton net.
3.
Larispongia magdalenae Carrera, 1998, holotype CEGH-UNC 17365 from Lari
Creek, Puna region, vertical view of the complete specimen showing open
spongocoel and part of the external surface.
4.
Detail of the same specimen showing dermal hexactines surrounding major gaps.
5.
Incrassospongia ramis Carrera, 1996, holotype CEGH-UNC 9308, lateral view
of a complete specimen from San Juan Formation, Cerro La Chilca, San Juan
Precordillera.
6.
Nexospongia sillaensis Carrera, 1996, upper surface of the holotype from
San Juan Formation, Cerro La Chilca, San Juan Precordillera CEGH-UNC 3613.
7.
Chilcaia bimuralis Carrera, 1996, photomicrograph
showing the homogeneous arrangement of the sexiradiates in one side of the
paratype CEGH-UNC 9336. San Juan Formation, Cerro La Chilca, San Juan Precordillera.
8.
Rugospongia viejoensis Carrera, 1996. Side view of the holotype CEGH-UNC 9252, San Juan
Formation, CerroViejo, San Juan Precordillera.
which
there was an adequate nutrient supply. Preservation of associated algae
indicates that rocks in which they occur accumulated in the photic zone.
The
siliceous hexactins occur together with conodont faunas of the Pygodus–periodon
community typical for deeper slope environments around Laurentia (Mehl and
Lehnert, 1997) and the Precordillera.
Because
of their relatively plastic morphology, shapes of the sponges may be indicative
of their environments and are considered as good paleoenvironmental indicators.
For example, columnar or tubular forms as for example Archaeoscyphia, occur
where currents were moderately strong. The tubular form aids in “sucking”
water through the system, like effluents up and out a chimney (Rigby, 1983).
Beresi
and Rigby (1993) described four general morphotypes of sponges present in the
Precordillera assemblages: 1) columnar, cylindrical-shaped forms, 2) cup-vase
and funnel-shaped forms, 3) discoid to flattened, saucer-shaped forms, and 4)
pear- or hemispherical-shaped forms.
Three
distinct sponge biofacies zones have been distinguished in the Cambro-Ordovician
of the Argentine Precordillera by Mehl and Lehnert (1997):
1.
Slope facies with Hexactinellida.
2.
Outer or deeper platform with lithistid demosponges (Orthocladina) and
subordinate hexactinellid and octactinellid spicules.
3.
Reef and reef-mound of the eastern Precordillera: sponge-algal mound with
lithistids (e.g. Archaeoschypia); octactinellid spicules from strata
within the reef horizons; assemblages of polyactinellid and heteractinellid
Calcarea and soft demosponges (inter-reef basins or in tidal channels).
Similar
conclusions were obtained from the carbonate-clastic sequence of the Ponón
Trehue section, southern Mendoza Province (Beresi and Heredia, 2000). There,
associations of exclusively heteractinellid spicules (Eiffeliidae) are
restricted to allochthonous blocks of the shallow platform of the San Juan
Formation. In the outer platform and slope, autochthonous calcarenites and dark
limestones only contain hexactin spicules. The heteractinid sponges and isolated
octactine-based spicules, show a general restriction to the middle (biohermal
and biostromal horizons) and upper parts of the San Juan Formation platform and
with highly diverse faunas (Keller and Flugel, 1996) in the interval
Arenig/early Llanvirn. Besides, isolated octactine-based spicules have been
registered in allochthonous blocks derived of this platform, at the Ponón Trehué
section (Beresi and Heredia, 2000).
Recently,
Carrera (2001) has recognized seven main biofacies in the San Juan Formation: 1)
Archeoscyphia-Calathium-Girvanella biofacies 2) Archaeorthis biofacies
3) Leptellinid biofacies 4) stromatoporoid biofacies 5) Petroria-Paralenorthis
biofacies 6) Demosponge biofacies and 7) Nileid biofacies. In biofacies 1
and 6, sponges dominate the assemblages.
According
to this author, this pattern of highly variable biozone content and the absence
of recurrent biofacies may imply the absence of refuge areas possibly related to
the isolation of the Argentine Precordillera terrane during its drifting away
from Laurentia.
A
clear change in biofacies framework is verified since the late Arenig with the
development of stromatoporids (Carrera, 1997).
Final
remarks
The
Argentine Ordovician sponge faunas are found in three different settings: 1)
carbonate platform, 2) volcaniclastic sequence and 3) black shales facies.
1)
The Ordovician lithistid demosponge faunas from the Precordillera occur in
limestone deposited in open subtidal environments, on a stable carbonate
platform environment. A diverse, well preserved associated fauna is mainly
composed of brachiopods, gastropods, trilobites, nautiloids, bryozoans,
crinoids, bivalves and diverse algae.
On
the other hand, hexactinellid sponges are much less common and diverse in the
Ordovician Precordillera, although isolated hexactin spicules have been
recognized in deeper environments from slope sequences.
Calcareous
heteractinida have a limited record. Heteractinid calcareous sponges are
represented by Chilcaia bimuralis Carrera and by the genera Dodecactinella,
Sardospongia and Eiffelia. The heteractinid sponges and isolated
octactine-based spicules, show a general restriction to the middle (biohermal
and biostromal horizons) and upper parts of the San Juan Formation in the
interval Arenig/early Llanvirn. Besides, isolated octactine-based spicules have
been recorded in allochthonous blocks at the Ponón Trehué section, Sierra
Pintada.
2)
The single hexactinellid sponge (Pelicaspongiidae) from the Puna region, western
margin of Gondwana, was found in fossiliferous sandstones of the Tremadocian
volcaniclastic sequence of the Las Vicuñas Formation. The associated fauna is
represented by trilobites and brachiopods.
3)
The material from the Famatina System is associated with deep environments. The
sponge material from the Lower Ordovician Volcancito Formation is preserved in
thinly laminated black siltstones and is associated with dendroids graptolites.
This black shales facies represents sedimentation from suspension fall-out in
very low-energy settings below storm wave base. The hexactinellid sponges of the
Famatina region occur in less favorable conditions that those of shelf
depositional environments. The sediments in which the sponges occurr suggest
restricted water circulation on the shelf, with oxygen depletion in the sea
floor (Esteban and Rigby, 1998). These protosponges document the occurrence of
such relatively primitive haxactinellid sponge in Paleozoic black-shale facies
at margins of the continents (Rigby et al., 1996; Esteban and Rigby,
1998).
Taxonomic
list of the new genera and species described from the Ordovician of Argentina
DEMOSPONGEA
-
Familia Anthaspidellidae Miller, 1889
Anthaspidella
alveola Beresi
and Rigby, 1993
Anthaspidella
annulata Beresi
and Rigby, 1993
Anthaspidella
inornata Beresi
and Rigby, 1993
Archaeoscyphia
nana Beresi and Rigby, 1993
Aulocopium
sanjuanensis Beresi and Rigby, 1993
Calycocoelia
perforata Beresi
and Rigby, 1993
Hudsonospongia
talacastensis Beresi
and Rigby, 1993
Incrassospongia
ramis Carrera,
1996
Nexospongia
sillaensis Carrera,
1996
Patellispongia
argentina Carrera,
1994
Patellispongia
inornata Beresi and Rigby, 1993
Patellispongia
robusta Beresi and Rigby, 1993
Psarodictyum
magna Beresi and Rigby, 1993
Rhopalocoelia
rama Beresi and Rigby, 1993
Rhopalocoelia
tenuis Carrera
1997
Rugospongia
viejoensis Carrera,
1996
Talacastonia
chela Beresi and
Rigby, 1993
CALCAREA
-
Familia Astraeospongiidae Miller, 1889
Chilcaia
bimuralis Carrera, 1994
HEXACTINELLIDA
-
Familia
Pelicaspongiidae Rigby, 1970
Larispongia
magdalenae Carrera,
1998
-
Familia Protospongiidae Hinde, 1887
Protospongia
sp. A Rigby and
Esteban, 1998
Protospongia sp. B. Rigby and Esteban, 1998
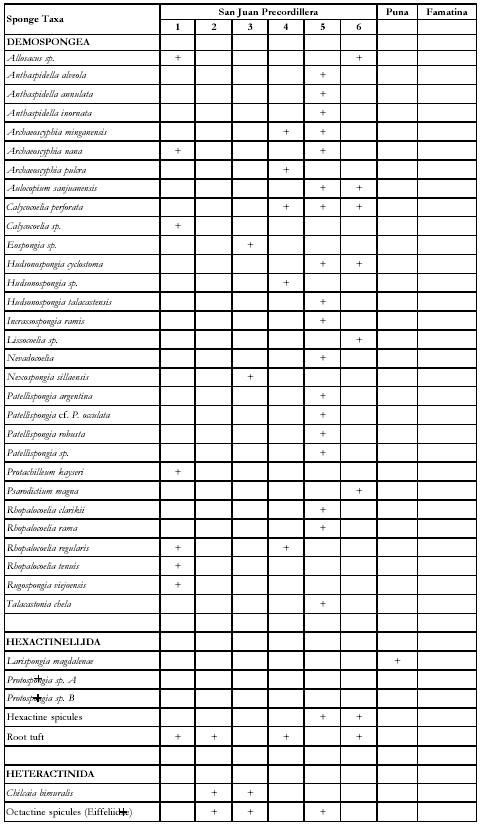
Table
1. Distribution of Ordovician sponges into Argentine
basins. Fossiliferous
localities: Huaco (1), Cerro La Silla (2), Cerro La Chilca (3), Las
Chacritas-Tunas sections (4), Talacasto Gulch (5), Villicum Range (6). Present +
References
Aceñolaza,
F.G. y Toselli, A.J. 1977. Observaciones geológicas y paleontológicas sobre el
Ordovícico de la zona de Chaschuil, Provincia de Catamarca. Acta Geológica
Lilloana, 14: 55-81. Tucumán.
Beresi,
M. 1985. Capas con Archaeoscyphia (Porífera) en los sedimentos carbonáticos
ordovícicos de la Precordillera de San Juan. 1º Jornadas sobre Geología de
la Precordillera, Actas, 1: 99-102. San Juan.
Beresi,
M. 1986. Paleoecología y Biofacies de la Formación San Juan, al sur del
paralello de 36º sur, Precordillera de San Juan. Tesis Doctoral, Universidad
Nacional de San Juan. Inédito. 450 pp.
Beresi,
M. 1990. Las esponjas como indicadores paleoecológicos de la Formación San
Juan, Precordillera. V Congreso Argentino de Paleontología y Bioestratigrafía,
Actas, 1: 19-24. Tucumán.
Beresi,
M. and Heredia, S. 2000. Sponge spicule assemblages from the Middle Ordovician
of Ponón Trehué, southern Mendoza, Argentina. Revista Española de Paleontología,
15 (1): 37-48.
Beresi,
M. and Rigby, J. K. 1993. The Lower Ordovician sponges of San Juan, Argentina. Brigham
Young University Geology Studies, 39: 1-64. Utah.
Cañas,
F. and Carrera, M.G. 1993. Early Ordovician microbial-sponge-receptaculitid
bioherms of the Precordillera basin, Western Argentina. Facies
29:169-178.
Erlangen.
Carrera,
M.G. 1985. Descripción de algunos poríferos de la Formación San Juan (Ordovícico)
de la Precordillera Argentina. Reunión de Comunicaciones Paleontológicas de
la Asociación Paleontológica Argentina, Resúmenes: 51-53.
Carrera,
M.G. 1994a. An
Ordovician sponge fauna from San Juan Formation, Precordillera basin, Western
Argentina. Neues Jahrbuch für Geologie und Paläontologie, 191 (2):
201-220. Abhandlunge.
Carrera,
M.G. 1994b. Taxonomía, Bioestratigrafía y significado paleoambiental de los
poríferos y briozoos del Ordovícico de la Precordillera Argentina. Tesis
Doctoral, Universidad Nacional de Córdoba. Inédito. 292 pp.
Carrera,
M.G. 1996a. Ordovician
Megamorinid sponges from San Juan Formation, Precordillera, Western Argentina. Geobios,
29(6): 643-650. Lyon
Carrera,
M.G. 1996b. Nuevos poríferos de la Formación San Juan (Ordovícico),
Precordillera Argentina. Ameghiniana, 33 (3): 335-342. Buenos Aires.
Carrera,
M.G. 1997a. Análisis paleoecológico de la fauna de poríferos del Llanvirniano
tardío de la Precordillera argentina. Ameghiniana, 34 (3): 309-316.
Buenos Aires.
Carrera,
M.G. 1997b. Evolución y recambio de las faunas de poríferos y briozoos en el
Ordovício de la Precordillera Argentina. Ameghiniana, 34 (3): 295-308.
Buenos Aires.
Carrera,
M.G. 1998. First
ordovician sponge from the Puna region, northwestern Argentina. Ameghiniana,
35 (2): 205-210.
Buenos Aires.
Carrera,
M.G. 2000. Distribución y composición de las biofacies de la Formación San
Juan (Ordovícico) Precordillera argentina. Ameghiniana 37, Suplemento
7R-8R. Buenos Aires.
Carrera,
M.G. 2001. Análisis de la distribución y composición de las biofacies de la
Formación San Juan (Ordovícico temprano), Precordillera argentina. Ameghiniana,
38 (2): 169-184. Buenos Aires.
Carrera,
M.G. and Rigby, J.K. 1999. Biogeography of the Ordovician sponges. Journal of
Paleontology, 73: 26-37.
Esteban,
S.B. and Gutiérrez-Marco, J.C. 1997. Graptolitos del Tremadoc del Sistema de
Famatina (Argentina). V Reunión Internacional Proyecto 351 PICG
“Paleozoico Inferior del Noroeste de Gondwana”, Libro de Resúmenes,
59-63. La Coruña.
Esteban,
S.B. and Rigby, J.K. 1998. Hexactinellid sponges from the Lower Tremadocian
Volcancito Formation, Famatina Range, Northwestern Argentina. Brigham Young
University Geology Studies, 43: 1-7. Utah.
Gnoli,
M. and Serpagli, E. 1980. A sponge spicula assemblage from Lower Ordovician of
Precordilleran Argentina. Rivista Italiana di Paleontología, 86: 267-272.
Heredia,
S. E. 1996. El Ordovícico
del Arroyo Ponón Trehué, sur de la Provincia de Mendoza, Argentina. XIII
Congreso Geológico Argentino, Actas, 1: 601-605. Buenos Aires.
Heredia,
S.E. 2001. Late
Llanvirnian conodonts from the Ponón Trehué Formation, Mendoza, Argentina. GAIA,
16 : 101-117. Lisboa.
Hünicken,
M. 1985. Lower Ordovician Conodont Biostratigraphy in Argentina. Boletín
de la Academia Nacional de Ciencias, Córdoba, 56
(3-4): 309-322.
Hünicken,
M.A. and Ortega, G. 1987. Lower Llanvirn-Lower Caradoc (Ordovician) conodonts and
graptolites from the Argentine Precordillera. In Austin, R.L. (ed.).
Conodonts: Investigative Techniques and Applications. 7:136-145
Kayser,
E. 1925. Contribuciones a la Paleontología de la República Argentina: Sobre fósiles
primordiales e infrasilurianos. Actas de la Academia Nacional de Ciencias,
Córdoba, 8 (3-4): 297-333.
Keller,
M. and Bordonaro, O. 1993. Arrecifes de estromatoporoideos en el Ordovícico inferior del Oeste
Argentino y sus implicancias paleogeográficas. Revista Española de
Paleontología, 8 (2): 165-169.
Keller,
M. and Flugel, E. 1996. Early Ordovician reefs from Argentina: Stromatopotroid
vs. Stromatolite origen. Facies, 34:
177-192.
Lemos,
V.B. 1981. Conodontes ordovicianos de Formaçao San Juan, Precordillera,
Departamento Jáchal, Provincia de San Juan, República Argentina
(Bioestratigrafía). Anais II Congresso Latinamericano de Paleontología,
1: 31-43. Porto
Alegre.
Mehl,
D. and Lehnert, O. 1997. Cambro-Ordovician sponge spicules assemblages in the
Ordovician of the Argentine Precordillera and paleoenvironmental. Neues
Jahrbuch für Geologie Paläontologie, 204:
204-246. Abhandlungen.
Peralta
S. P.; Heredia S. y Beresi, M. S. 1999. Estratigrafia del Ordovícico del Río
de Las Chacritas, sierra de la Trampa, Precordillera central de San Juan,
Argentina. 8th
International Symposium on the Ordovician System. Acta
Carolinae, 43
(1/2): 123-127. Czech Republic.
Sarmiento,
G. N. 1985. La biozona de Amorphognathus variabilis-Eoplacognathus
pseudoplanus (Conodonta), Llanvirniano inferior, en el flanco oriental de la
Sierra de Villicúm. 1º Jornadas sobre Geología de la Precordillera, Actas 1:
119-123. San Juan.
Rigby,
J. K. 1983. Introduction
to the Porifera. In T.W. Broadhead (ed), Sponges and Spongiomorphs,
University of Tennessee, 201pp.
Rigby,
J. K.; Gutiérrez-Marco, J.C.; Robardet, M. and Picarra, J.M. 1996. First
articulated Silurian sponges from the Iberian Penninsula, Spain and Portugal. Journal
of Paleontology, 71: 554-563.
Turner,
J.C. 1964. Descripción
geológica de la Hoja 15c, Vinchina (provincia de La Rioja). Boletín de la
Dirección Nacional de Geología y Minería, 100: 81pp.
Recibido:
4 de Septiembre
de 2002