Icnofacies Skolithos-Ophiomorpha en el Neogeno del valle inferior del río Chubut, provincia del Chubut, Argentina
Roberto R. LECH (1) , Florencio G. ACEÑOLAZA (2) y Mercedes M. GRIZINIK (3) .
Abstract.- THE ICHNOFACIES SKOLITHOS-OPHIOMORPHA IN NEOGENE OF THE LOWER VALLEY OF RIO CHUBUT, CHUBUT PROVINCE, ARGENTINA. The icnocoenosis Skolithos-Ophiomorpha of marine Neogen strata outcroping in lower valley of "Río Chubut", province of Chubut are described. The geological sections have common Ophiomorpha nodosa and the icnocoenosis of Cylindrichnus concentricus, Planolites beverleyensis, Polykladichnus irregularis, Rosselia socialis, Skolithos linearis y Thalassinoides suevicus.
The association is representative of one littoral zone, intertidal to subtidal, with notable continuity along the thickness. Same confirm previous ideas on only one marine event in Middle Miocene (Tortonian) times.
Key words: Trace fossils. Miocene. Patagonia
Palabras Claves: Icnofósiles. Mioceno. Patagonia.
Introducción
Recientes trabajos sobre el Neógeno marino aflorante en el tramo inferior del valle del río Chubut nos permitieron reconocer que un importante espesor sedimentario se encuentra intensamente bioturbado con la típica asociación icnológica integrada por Skolithos-Ophiomorpha.
La sola mención de dichas icnoespecies en la columna estratigráfica que caracteriza a las Formaciones Gaiman y Puerto Madryn, nos permite señalar algunos aspectos interesantes sobre la biodiversidad de las misma y a la vez replantear conceptos estratigráficos que apuntalan las nuevas ideas que se tiene sobre las fluctuaciones del nivel del mar en la costa atlántica durante el Terciario medio (Tortoniano).
Debe reconocerse que durante mucho tiempo la discusión sobre la posibilidad de estar ante una serie de eventos transgresivos-regresivo estuvieron atados a interpretaciones tectogénicas y a las determinaciones de edades relativas basadas en la malacofauna fósil presente en los niveles marinos. Hoy los numerosos estudios desarrollados sobre la edad de la fauna, como asimismo, los llevados a cabo por Haq et al. (1988) sobre los registros mundiales de los cambios del nivel del mar; como asimismo a las revisiones de la fauna de moluscos llevada adelante por Del Río et al. (1999) contribuyen a mejorar la interpretación que se tiene sobre las unidades marinas del Neogeno patagónico.
El material fósil estudiado nos permite reconocer en las unidades estratigráficas asignadas a Gaiman y Puerto Madryn una secuencia, en términos generales, icnologicamente continua, típica de la región intermareal y submareal en los ambientes marino de transición. Las discontinuidades icnológicas observadas en el campo puede deberse tanto a cambios de corta duración en la energía del ambiente sedimentario como a variaciones de la línea de costa originada por eventos eustáticos diferentes.
Antecedentes
Los primeros estudios geológicos sobre el Terciario de Patagonia fueron realizados por d´Orbigny (1842) y Darwin (1846), quienes al realizar una reseña de los afloramientos del litoral atlántico respectivamente los describen como "Tertiaire patagonién" y "Patagonian Tertiary Formation".
Debe destacarse que las descripciones más detalladas fueron desarrolladas, años más tarde, por Bravard (1858), F. Ameghino (1889, 1894, 1897, 1898 y 1906), Burmeister (1876), Döering (1882), C. Ameghino (1890), Ortman (1902), Wilckens (1905), Ihering (1907), Roveretto (1921), Windhausen (1919, 1921), Frenguelli (1926), Feruglio (1949, 1950) entre otros, quienes se involucran en discusiones sobre el desarrollo de los estratos marinos y continentales presentes en distintos sectores de la costa atlántica.
La definición de varios ciclos transgresivos-regresivos durante el Terciario inferior y medio llevaron a la instalación, en la nomenclatura geológica, de una serie de nombres cuya interpretación cronológica y estratigráfica dio origen a interpretaciones muchas veces controversiales. Así por ejemplo nombres como "Patagoniense", "Suprapatagoniense" "Entrerriense", "Leonense", "Camaronense", "Aonikense" o "Juliense" han sido objeto de diversas consideraciones, no siempre acordes con el hoy vigente Código de Nomenclatura Estratigráfica.
Por las características del presente trabajo no entraremos en detalles sobre el debate mencionado, aunque debemos señalar que, si bien el mismo aportó más detalles a las descripciones estratigráficas y paleontológicas, también en muchos casos oscureció el panorama interpretativo.
Sin perjuicio de ello debemos destacar que Frenguelli (1926, 1935) realizó un detallado relevamiento de la estratigrafía de la zona de Gaiman, reconociendo que sobre la «serie tobífera» (Formación Sarmiento) es factible observar una sucesión integrada por el «Patagoniense», «Aonikense» (sensu Rovereto, 1921) y el «Entrerriense». En este conjunto Haller y Mendía (1980) interpretan que la secuencia neogena puede integrarse en las Formaciones Gaiman y Puerto Madryn, esta última portadora de la malacofauna caracterizada por Ostrea madryna. En cambio Scasso y Del Río (1987), luego de un exhaustivo estudio de la fauna y sedimentología llegan a la conclusión que tanto el «Patagoniense», «Entrerriense» y «Rionegrense» constituyen un único evento sedimentario marino de carácter regresivo, criterio que también hemos sostenido en una comunicación sobre este tema (Aceñolaza et al, 1998).
Marco geológico
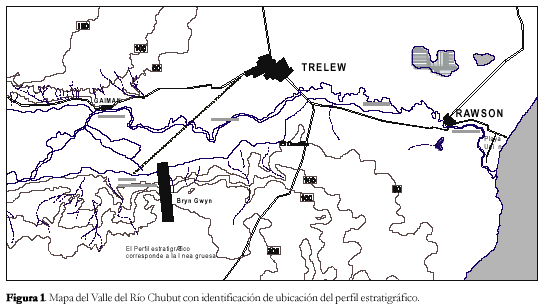
La geología del sector conocido como Lomas Blancas o «Bryn Gwyn» (en idioma Galés), al sur del río Chubut, fue objeto de reiterados trabajos tanto estratigráficos como paleontológicos, debido a la importancia que se le atribuye en la interpretación evolutiva del Terciario aflorante en el valle inferior del río Chubut. Como ya hemos mencionado, Frenguelli (1935) es quien tal vez mayor atención le prestara a la columna allí aflorante describiéndola en sus detalles centimétricos. Años más tarde, Feruglio (1949, 1950) y recientemente Franchi et al. (1975), Haller y Mendía (1980), Mendía y Bayarsky (1981) y Scasso y Castro (1999) se abocaron con mayor detenimiento al estudio del sector mencionado aportando distintas interpretaciones sobre ambos pisos. Especialmente Haller y Mendía (1980) describieron en la secuencia marina allí aflorante a las unidades formacionales de Gaiman y Puerto Madryn.
Para estos últimos autores la Formación Gaiman tiene su localidad tipo en Bryn Gwyn (Bardas Blancas), al sur del río Chubut frente a la localidad de Gaiman. El espesor máximo es del orden de los 140 metros, estando constituida por una sucesión de areno-arcillas con material predominantemente tobáceo, de colores muy claros con intercalaciones con restos fósiles de invertebrados marinos. Su relación de base es discordante sobre las tobas con mamíferos terrestres de la Formación Sarmiento.
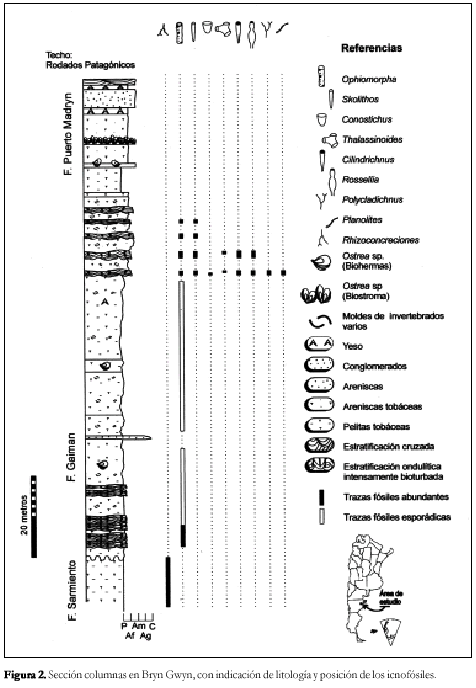
Desde el punto de vista sedimentológico, el contacto entre las mencionadas Formaciones es difícil de observar, debido a que los sedimentos de la base la Formación Gaiman provendrían de la remoción por acción del agua de mar de aquellas sedimentitas que constituyen a la Formación Sarmiento. Sin embargo, esta discordancia esta limitada por trazas fósiles de distinta naturales que nos permiten inferir su presencia. En los términos superior de la Formación Sarmiento las rhizoconcreciones son abundantes y desaparecen rápidamente hacia el contacto con la Formación Gaiman. En cambio en la base de esta última Formación se encuentran las primeras Ophiomorpha que caracterizaran gran parte de la secuencia sedimentaria marina miocena del valle inferior del río Chubut.
La Formación Puerto Madryn fue propuesta por Haller (1979) para las psamitas y pelitas de tonalidades más verdosas que aquellas que caracterizan a la Formación Gaiman, y se extienden por Península Valdés, el Golfo Nuevo, y el valle inferior del río Chubut (Feruglio, 1949, 1950).
Frente a las localidades de Trelew y Gaiman esta unidad estratigráfica presenta un espesor sedimentario del orden de los 30 metros, integradas en una secuencia de arenas, limolitas y arcilitas que están fuertemente bioturbadas con distintos elementos representativos de la icnocoenosis Skolithos-Ophiomorpha. Debe destacarse que el paso de la Formación Gaiman a la Formación Puerto Madryn es normal, pudiendo notarse que los icnofósiles tienen una continuidad vertical notable de una a otra.
En la Figura 1 se representa las características sedimentológicas y la distribución estratigráfica de los icnofósiles descritos para las Formaciones Gaiman y Puerto Madryn (sensu Haller y Mendía, 1980) aflorante en las bardas sur frente a la localidad de Gaiman.
Descripción icnológica
El material fósil descripto a continuación procede principalmente de las secuencias sedimentarias aflorantes en Bryn Gwyn, Gaiman, además de Loma del Castillo, Trelew, y Playa Maganea, al sur de Playa Unión, Rawson.
Icnogénero Cilyndrichnus Howard, 1966
Cilyndrichnus concentricus Howard, 1966
Material: Numerosos ejemplares «in locus» en la Formación Puerto Madryn, aflorante en Bryn Gwyn, Gaiman.
Descripción: Estructura tubiforme subcilíndrica recta o ligeramente curvada, dispuesta perpendicular al plano de estratificación. Las paredes están constituidas por capas concéntricas de sedimento, que se expanden ligeramente en la posición oral.
Observación: En secciones transversas del material asignado a esta icnoespecie se observa las estructuras concéntricas de las capas que constituyen la pared. El diámetro máximo medido en la zona de expansión entre los distintos ejemplares oscila entre los 15 y 20 mm.
Materiales de este tipo han sido descriptas por Frey y Howard (1985) para el Cretácico de Estados de Unidos y para el Mioceno (Formación Paraná) de Entre Ríos, Argentina (Aceñolaza y Aceñolaza, 2000) Icnogénero Conostichus Lesquereux, 1876 Conostichus isp.
Figura 3, b
Material: Numerosos ejemplares «in locus» en la formación Puerto Madryn aflorantes en Bryn Gwyn, Gaiman.
Descripción: Estructuras cónicas verticales, ornamentadas externamente por crestas y constricciones. Las crestas y surcos se disponen longitudinalmente y pueden superponerse. Poseen disco apical hemisférico. La altura esta en el orden de los 5 cm y el diámetro máximo llega a 7 cm. En un corte transversal se observa la estructura interna constituida por un relleno con forma de meniscos cóncavos hacia arriba.
Observaciones: Conostichus se encuentra generalmente en secuencias de plataforma marina.
En nuestro país este icnogénero es conocido para el Cámbrico (Aceñolaza et al., 1982) y el Carbonífero (Lech y Buatois, 1990).
Icnogénero Ophiomorpha Lundgren, 1891 Ophiomorpha nodosa Lundgren, 1891.
Figura 3, a, c y c’, Figura 3, a y Figura 4, d
Material: Numerosos ejemplares «in locus» en las Formaciones Gaiman y Puerto Madryn aflorantes en Bryn Gwyn, Gaiman; en la Formación Gaiman aflorante en Loma del Castillo, Trelew, y en la Playa Maganea, al sur del puerto de Rawson.
Descripción: Sistema de galerías rectas, verticales y horizontales, de sección circular con un diámetro variable entre unos pocos centímetros y 16 centímetros máximo. Las galerías horizontales se ramifican irregularmente tanto en forma de Y como de T. La longitud vertical visible de las distintas galerías oscila en el orden de los decímetros. Exteriormente las paredes están cubiertas por gránulos subesféricos irregulares que pueden llegar a 16 mm de diámetro máximo en las galerías de mayor tamaño, en tanto que el interior de las galerías es liso.
Observaciones: Esta icnoespecie se la encuentra en la parte alta de la barda de Bryn Gwyn forman un intrincado sistema de galerías verticales y horizontales, de variado diámetro y gran extensión horizontal. Se preservada generalmente como endichnia tanto en sedimentos carbonáticos como silíceos y es especialmente abundantes en rocas de edad mesozoica y cenozoica (Pemberton y Jones, 1988). Ha sido interpretado como galerías de alimentación (Frey et al., 1978) o combinada de alimentación-habitación (Frey y Howard, 1985) y se las atribuye como originada posiblemente por crustáceos decápodos semejantes al grupo de los talasínidos.
Actualmente se reconocen cuatro icnoespecies diferenciadas por la forma de las paredes, las mismas son: O. annaluta, O. borneensis, O. irregulaire y O. nodosa (Pemberton y Jones, 1988). Debemos hacer notar que Scasso y Castro (1999) hace mención e ilustra ejemplares de esta icnoespecie procedentes del mismo sector en estudio.
Icnogénero Planolites Nicholson, 1873
Planolites beverleyensis (Billings, 1862)
Figura 3, e, Figura 4, c y f
Material: Numerosos ejemplares «in locus» en la Formación Puerto Madryn, Bryn Gwyn, Gaiman.
Descripción: Estructuras tubiformes dispuesta paralelamente al plano de estratificación, rectas a meandriformes, raramente ramificadas. El diámetro de los tubos oscila alrededor de los 5 mm, si bien es posible encontrar secciones irregulares de diámetro mayor. La sección puede ser circular a elíptica. No se observó estructuras sedimentarias en el interior de las galerías, y el sedimento difiere en color con respecto a la roca de caja.
Observaciones: Pemberton y Frey (1982) desarrollan en un extenso trabajo cual es la característica principal del icnogénero Planolites y sus diferencias con Palaeophychus con quien es posible confundir. Señalan así también, que las principales diferencias entre ambos icnogéneros corresponden al hecho que el relleno de Planolites es diferente en estructura y composición respecto al material de la roca de caja. Los mismo autores, basándose en el tamaño, curvatura y características morfológicas de la pared de Planolites, reconocen como válidas a tres icnoespecies: P. montanus, P. beverleyensis y P. annularis.
P. beverleyensis se diferencia de las otras dos especies por el tamaño, por la configuración recta a curva de las galerías y por las paredes lisas (Pemberton y Frey, 1982). Esta traza fósil se preserva generalmente como endichnia, hipicnia y epicnia, y ha sido interpretada como una estructura de alimentación (Frey y Howard, 1985).
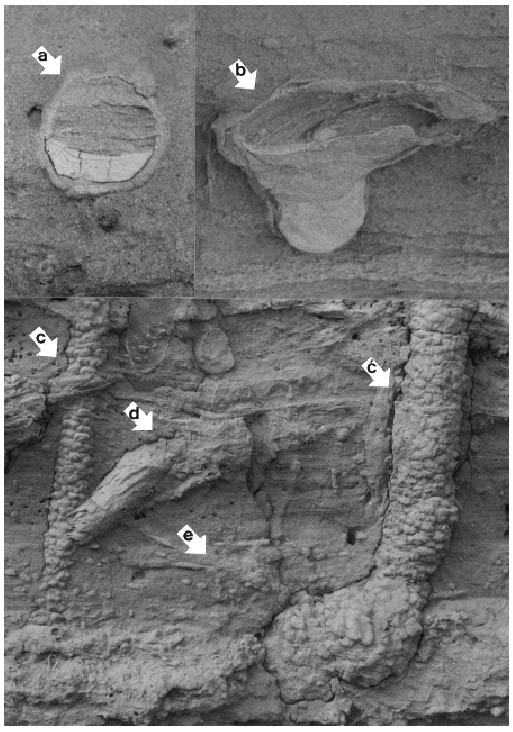
Figura 3. a: Corte transversal de un tubo horizontal de Ophiomorpha nodosa. b: Conostichus isp. c y c´: Tubos verticales de Ophiomorpha nodosa donde se observa el ensanchamiento de la base formando un nodo al horizontalizarse el tubo. d: Rosselia socialis removida de su posición normal. e: Nivel con Planolites beverleyensis. Todo el material fósil fotografiado proviene de la Formación Puerto Madryn, Bryn Gwyn, Gaiman.
Planolites es un icnogénero con un rango estratigráfico muy amplio, que va desde el Precámbrico superior hasta la actualidad. Hay autores que lo atribuyen como originado por anélidos poliquetos del grupo de los eunícidos (Howard y Frey, 1975), o bien por hemicordados del tipo de Balanoglossus (Rupper y Barnes, 1996).
Icnogénero Polykladichnus Fürsich, 1981 Polykladichnus irregularis Fürsich, 1981 Figura 5, a y a’
Material: Varios ejemplares observados «in locus» en la Formación Puerto Madryn, Bryn Gwyn, Gaiman.
Descripción: Trazas tubulares rectas o ligeramente curvas, bifurcadas en Y, perpendiculares al plano de estratificación y en general conectadas con la superficie de sedimentación. El diámetro oscila entre los 4 y 8 mm y la longitud máxima observada alcanza los 17 cm. No se observó estructuras internas, y cuando falta el relleno se observa las paredes constituidas por granos de arena cementados por carbonatos.
Observaciones: Polykladichnus si bien es poco frecuente, se identifica perfectamente por su particular forma de Y, acompañando a los niveles con Skolithos, forma con quien puede confundirse cuando falta alguna de las ramas o cuando está se verticaliza demasiado. Pemberton y Jones (1988) destacan la abundancia de Polycladichnus en sedimentos del Pleistoceno de las islas Gran Cayman acompañando los niveles con Skolithos. Así mismo Polycladichnus a sido mencionado por Fürsich (1981) en el Jurásico de Portugal, interpretandolo que su origen se debió a la actividad de poliquetos marinos.
Icnogénero Rosselia Dahmer, 1937
Rosselia socialis Dahmer, 1937.
Figura 3, d, Figura 5, e
Material: Numerosos ejemplares «in locus» en la Formación Puerto Madryn, Bryn Gwyn, Gaiman, y en la Formación Gaiman aflorante en Loma del Castillo, Trelew.
Descripción: Estructura tubular de aspecto general cónica invertida con expansiones irregulares, que se dispone en posición vertical o inclinada con respecto al plano de estratificación.
La longitud máxima medida en los ejemplares reconocidos es de 30 cm, en tanto que el diámetro máximo en igual ejemplar es de 5,5 cm. El extremo oral se cierra ligeramente, otorgándole un aspecto globoso; a medida que la traza crece puede presentarse una segunda expansión globosa, de mayor tamaño que la anterior.
A pesar de que en el material observado no se preservó el extremo distal, el mismo aparenta terminar en un tubo de sección circular de alrededor de un centímetro de diámetro constituido por tres capas de sedimento de distinta granulometría fina. En el extremo oral, siete capas concéntricas de sedimento fino a muy fino forman al cuerpo; así también un cilindro formado por arena fina recorre interiormente y a lo largo de toda su longitud a la estructura de bioturbación.

Observaciones: Rosselia puede ser confundido algunas veces tanto con Cylindrichnus, Skolithos, y Asterosoma (Frey y Howard, 1985), y ha sido interpretado como galerías de alimentación de anélidos filtradores de sedimento (Pemberton et al., 1992).
En sucesiones sedimentarias del Cretácico Rosselia y Asterosoma son excelentes indicadores de ambientes marinos de plataforma afectados por las olas de tormentas (Pemberton et al. , 1992).
Icnogénero Skolithos Haldeman, 1840 Skolithos linearis Haldeman, 1840
Figura 4, b y b’, Figura 5, b
Materiales: Numerosos ejemplares «in locus» en la Formación Puerto Madryn, Bryn Gwyn, Gaiman, y en la Formación Gaiman aflorante en Loma del Castillo, Trelew, y en la Playa Maganea, al sur del Puerto de Rawson.
Descripción: Traza tubiforme vertical a subvertical, cilíndricas a subcilíndricas, de paredes lisas. El diámetro máximo se ubica en el extremo oral, y oscila alrededor de 1 cm, en tanto que la longitud máxima no supera los 10 cm. El relleno no presenta estructuras aparentes y el sedimento contrasta ligeramente con la roca de caja.
Observaciones: Debemos destacar que las asignaciones específicas en el icnogénero Skolithos se tornan dificultosas debido a los escasos elementos morfológicos de éste presenta, esto motiva que las asignaciones icnoespecíficas sean numerosas y confunden el panorama general respecto a su taxonomía. Tomando en consideración la revisión sistemática de Alpert (1974) y los comentarios taxonómicos de Fillion y Pickeril (1990), se reconocen actualmente como válidas solo siete icnoespecies de Skolithos: S. linearis; S. verticalis; S. serratus; S. magnus; S. ingens; S. annulatus; S. bulbus, y S. gyratus.
Las características morfológicas descriptas para los ejemplares de Skolithos de Bryn Gwyn (posición vertical, sección cilíndrica a subcilíndrica, paredes lisas) corresponden a aquella típica traza vertical y recta correspondientes a la icnoespecie S. linearis. Skolithos es una traza de tipo domicnia encontrada en variados ambientes de depositación, tanto marinos someros a profundos (Ranger y Pemberton, 1988; Hallam y Swett, 1966; Crimes, 1977) como no marinos (Filzgerald y Barret, 1986). En tanto que el registro paleontológico abarca desde el Precámbrico al Pleistoceno (Fedonkin, 1985; Pemberton y Jones, 1988).
Icnogénero Thalassinoides Ehrenberg, 1944
Thalassinoides suevicus (Rieth, 1932)
Materiales: Abundantes ejemplares «in locus» en la Formación Puerto Madryn, Bryn Gwyn, Gaiman.
Descripción: Estructuras tubiformes ramificadas, curvas y de sección cilíndrica a subcilíndrica, con el diámetro variable entre 5 y 10 mm. Las mismas forman un sistema de galerías paralelas al plano de estratificación o las interceptan ligeramente. No se ha detectado la

pared, y la superficie externa de las galerías es lisa. El relleno no presenta estructuras y el color del mismo contrasta ligeramente con el sedimento de caja.
Observaciones: Thalassinoides es una traza del tipo epicnia, endicnia o hipicnia, originada por organismos detritívoros afines a los crustáceos decápodos del grupo de los callasínidos (Howard y Frey, 1984; Bromley, 1967). Ha sido interpretada como una traza de alimentación-habitación, característica en sedimentos finos y coherentes, donde el refuerzo de las paredes es innecesario (Frey y Howard, 1985).
Este icnogénero ha sido registrado a partir del Ordovícico (Aceñolaza y Fernández, 1984; Sheenan y Schiefelbein, 1984) y es muy abundante a partir del Mesozoico y Cenozoico. Se lo encuentra en secuencias sedimentarias de origen marino somero (Howard y Frey, 1984), abanicos submarinos profundos (Crimes, 1977; Buatois y López Angriman, 1991) y en secuencias marinas transicionales «shoreface-offshore» (Mángano y Buatois, 1994).
Actualmente se reconocen dos icnoespecies de: T. paradoxicus y T. suevicus. La primera presenta una alta variabilidad en sus sistemas, una ramificación irregular y por cortar marcadamente los planos de estratificación (Fürsich, 1974).
Consideraciones estratigráficas y paleoambientales
En una comunicación anterior (Aceñolaza et al., 1998), teniendo en cuenta la icnofauna presente en el perfil de Bryn Gwyn y en los alrededores de Gaiman, habían comprobado que la columna estratigráfica del Neogeno marino allí aflorante tenía una notable continuidad vertical.
Esta situación nos llevó a pensar que la diferenciación en Formaciones de alguna manera perdía sustento, criterio que nos acercó a la posición de Scasso y Del Río (1987) y Scasso y Castro (1999). La continuidad vertical de las icnoespecies, particularmente las de Ophiomorpha nodosa, nos indicaban que se estaba ante un único evento en el cual ambos términos estratigráficos representaban oscilaciones en un ámbito litoral. O sea que la visible variación cromática, y eventualmente litológica, más se debía a las características del material que las originó que a eventos eustáticos diferentes. Dicho de otra forma, la Formación Gaiman (sensu Haller y Mendía, 1980) debía sus caracteres a que su composición representaba la remoción y reciclaje de la Formación Sarmiento; mientras que en la Formación Puerto Madryn solo se acentuaban las características marinas. Este planteo nos llevó a sostener la conveniencia de que ambas sean nominadas como miembros de una única unidad estratigráfica.
Observando los caracteres bioestratigráficos de la secuencia se comprueba que la instalación del mar se inicia en una playa intermareal que pasa a submareal en los niveles portadores de Ostrea máxima. El carácter transgresivo no aparece reflejado con claridad en la litología, pero si por la presencia de niveles con malacofauna y abundantes restos fósiles óseos de vertebrados marinos y dientes de tiburones. En esta unidad es notoria la presencia de Ophiomorpha que, como se conoce, es representativa de zonas intermareales hasta submareales. Se interpreta que esta estructura puede ser originada por el modo de vida de un grupo de decápodos, del grupo de los tallasínidos, cuya actividad sobre el bentos es mayor en la marea alta, mientras que en la baja la intrincada red de tubos les sirve de refugio y también para reproducción y desarrollar gran parte de su actividad vital.
De allí que la continuidad de este icnogénero a lo largo de la columna estratigráfica nos lleve a pensar que en ella solo se está ante una ligera fluctuación del nivel del mar o, dicho de otra manera, frente a variaciones de la línea de costa. Variaciones que, naturalmente, no debieron ser significativas. Debemos recordar que para Bellosi (1996) las grandes ostras que caracterizan a determinados niveles de la Formación Chenque, en Comodoro Rivadavia, no solo se ubican topográficamente en la zona litoral, sino que en muchos casos representan verdaderos biohermos originados en zona de manglares, detrás de la línea de mar. Interpretaciones de este tipo son concordantes con las de Scasso y Castro (1999) en el sentido que la unidad Gaiman se inicia en un frente de playa y aguas poco profundas con abundantes niveles de tempestitas para cerrar en niveles de tipo estuarino. La continuidad de las Ophiomorpha nos indica que nunca se retiró el mar ya que los niveles de Ostrea madrina del techo solo indican la instalación de un reflujo marino, por ejemplo una albúfera salobre, siempre en el ámbito litoral.
La icnocenosis, sigue siendo característica de zonas inter a submareales: Ophiomorpha, Thalassinoides, Skolithos y Rosselia son elementos visibles en determinados niveles. La continuidad lateral de las zonas bioturbadas es relativamente corta aunque a todas se las reconozca en posición estratigráfica variable. Este detalle es indicativo que la zona fue litoral y que como tal, estuvo sujeta a profundizaciones o somerizaciones en gran parte regidas por la dinámica costera.
La relativa abundancia de Skolithos y Ophiomorpha en facies de arenas finas, con una relativa buena selección indica que se está en ámbitos de playas abiertas en el frente marítimo; mientras que los Thalassinoides, normalmente presentes en bancos más arcillosos nos pueden indicar que estas condiciones han variado, por la presencia de dunas o barras que limitaron la energía de las aguas. Situaciones de este tipo se han comprobado en las trazas de la Formación Paraná, en el sentido que la paleogeografía, de alguna manera, condicionó la presencia de algunas icnoespecies (Aceñolaza y Aceñolaza, 2000).
Por último debemos señalar que la asociación de trazas nada nos dice sobre la cronoestratigrafía de la secuencia ya que, en su totalidad, son formas de amplio rango temporal.
Creemos que el debate debe hacerse con la abundante micro y megafauna presente en esta secuencia.
Bibliografia
Aceñolaza, F.G. y Aceñolaza G.F., 2000. Trazas fósiles del Terciario marino de Entre Ríos (Formación Paraná, Mioceno medio), República Argentina. Boletín de la Academia Nacional de Ciencias, 64: 209-233.
Aceñolaza, F.G. y Fernández, R.I., 1984. Nuevas Trazas Fósiles en el Paleozoico Inferior del Noroeste Argentino. Actas del 3º Congreso Argentino de Paleontología y Bioestratigrafía, 1: 13-28.
Aceñolaza, F.G., Fernández, R.I. y Manca, N., 1982. Caracteres bioestratigráficos y paleoambientales del Grupo Mesón (Cámbrico medio-superior), Centro-Oeste de América del Sur. Estudios Geológicos, 38: 385-392.
Aceñolaza, F.G., Lech, R.R. y Grizinik, M., 1998. Icnofacies Skolithos-Ophiomorpha en el Terciario marino del valle inferior del río Chubut. 3º Jornadas de Comunicaciones de la Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Resúmenes: 100.
Alpert, S.P., 1974. Systematic review of the genus Skolithos. Journal of Paleontology, 48: 661-669. Tulsa.
Ameghino, C., 1890. Exploraciones geológicas en Patagonia. Boletín del Instituto Geográfico Argentino, 11(1): 3-46.
Ameghino, F., 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Boletín de la Academia Nacional de Ciencias, 6: 1-1028.
Ameghino, F., 1894. Enumération synoptiques des espécès de mammiféres fossiles des formations éocenes de patagonie.
Boletín de la Academia Nacional de Ciencias, 13: 259-452 (1892-3).
Ameghino, F., 1897. Mammiféres crétaces de l´Argentine. Deuxime contribution a la connassaince de la faune mammologique des couches a Pyrotherium. Boletín del Instituto Geográfico Argentino. XVIII.
Ameghino, F., 1898. Sinopsis geológica-paleontológica. Segundo Censo de la República Argentina. 1. Buenos Aires.
Ameghino, F., 1906. Les formations sédimentaires du Crétacé superieur et du Tertiarie de Patagonie avec une parallele entre leurs faunes mammalogiques et celles de l´ancient continent. Annales del Museo Nacional de Historia Natural, 15 (3º serie): 1- 568.
Bellosi, S., 1996. Arrecifes de ostras gigantes en depósitos de albufera de la Formación Chenque (Mioceno Inferior, Cuenca San Jorge). 6º Reunión Argentina de Sedimentología Actas: 175-180. La Plata.
Billings, E., 1862. New species of fossil from different parts of the Lower, Middle, and Upper Silurian rocks of Canada. En: Palaeozoic Fossils. (1861 - 1865). Geological Survey of Canada, 1: 96-168. Canadá.
Bravard, A., 1858. Monografía de los terrenos marinos terciarios de las cercanías del Paraná. Impresión Registro Oficial, 107 pp. (Reimpresión Imprenta del Congreso de la Nación Argentina, 1995).
Bromley, R.G., 1967. Some observations on burrows of thallassinidean Crustacea in chalk hardgrounds. Geological Society of London, Cuarterly Journal, 123: 157-182.
Buatois, L.A. y López Angriman, A.O., 1992. The ichnology of a submarine braided channel complex: the Whisky Bay Formation, Cretaceous of James Ross Island, Antartic. Palaeogeography, Palaeoclimatology, Palaeoecology, 94: 119-140.
Burmeister, 1876. Description Phisique dela Republica Argentina,Climatolgie et Tableau géognostique, 2: 1-412. París.
Crimes, T.P., 1977. Trace fossils of an Eocene deeps-sea sand fan, northern Spain. En: Crimes, T.P. y Harper, J.C. (Eds.). Trace Fossils 2. Geological Journal, Special Issue 9: 71-90.
Dahmer, G., 1937. Lebensspuren aus dem Taunusquarzit udn den Siegener Schichten (Unterdevon). Preussischen Geologischen Landesanstalt zu Berlin, Jahrbuch 1936, 57: 523-539.
Darwin, Ch., 1846. Geological observations of coral reefs, volcanic islands and on South America.
Del Río, C.J., Martínez, S. y Scasso, R.A., 1999. Reconstrucción de las asociaciones de invertebrados bentónicos de la Formación Puerto Madryn (Mioceno) en Península Valdés (Provincia del Chubut). 14º Congreso Geológico Argentino, 1: 47. Salta.
Döering, A., 1882. Informe Oficial de la Comisión Científica agregada al estado mayor general de la expedición al Río Negro (Patagonia), Buenos Aires, Entrega 3, Geología.
Ehrenberg, K., 1944. Ergänzende Bemerkungen zu den seinerzeit aus dem Miozän von Burgschleinitz beschriebenen Gangeman und Bauten dekapoder Krebse. Paläontologische Zeitschrift, 23: 354-359.
Fedonkin, M.A., 1985. Paleoichnology of Vendian Metazoa. En B. Sokolov y M. Ivanovsky (Eds.) The Vendian System: Historic - geological and palaeontological basin. 1: 112-116.
Feruglio, E., 1949, 1950. Descripción Geológica de la Patagonia. Dirección General Yacimientos Petrolíferos Fiscales. Tomos I, II y III.
Filzgerald, P.G. y Barret, P., 1986. Skolithos in a Permian braided river deposit, southern Victoria Land. Antartica. Palaeogeography, Palaeoclimatology, Palaeoecology, 52: 237-247.
Fillion, D. y Pickerill, R.K., 1990. Ichnology of the Lower Ordovician Bell Island and Waban Group of Eastern Newfoundland. Palaeontographica Canadiana, 7. 1-119.
Franchi, M. Haller, M. , Lapido O.; Page, R. y Pesce, A. 1975 Geología de la región nororiental de la provincia de Chubut, República Argentina. Actas 2do Congreso Ibero-americano de Geología Económica, 4, 125-139.
Frenguelli, J., 1926. El Entrerriense del Golfo Nuevo en el Chubut. Boletín de la Academia Nacional de Ciencias, 29: 191-270. Córdoba.
Frenguelli, J., 1935. El perfil de Gaiman. Notas del Museo de La Plata, 1 (Geología, 1): 177-192.
Frey, R.W. y Howard, J.D., 1985. Trace fossils from the Panther Member, Star Point Formation (Upper Cretaceous), Coal Creek Canyon, Utha. Journal of Paleontology, 59 (2): 370-404.
Frey, R.W., Howard, J.D. y Pryor, W.A., 1978. Ophiomorpha: ist morphologic, taxonomic and environmental significance. Palaeoclimatology, Palaeogeography, Paleoecology, 23: 199-229. Amsterdam.
Fürsich, F.T., 1974. Ichnogenus Rhizocarallium. Paläntologische Zeitschrift, 48: 16-28.
Fürsich, F.T., 1981. Invertebrate trace fossils from the Upper Jurassic of Portugal. Comunicâcoes Servicos Geológicos de Portugal, 67: 1153-168.
Haldeman, S.S., 1840. Supplement to number one of "Amonograph of the Limniades, and other freshwater univalve shells of North America", containing descriptions of apparently new animals in different classes, and the names and characters of the subgenera in Paludina and Anculosa: 3p.
Hallam, A. y Swett, K., 1966. Trace fossils from the Lower Cambrian Pipe Rock of the north-west Highlands. Scottish Journal of Geology, 2: 101-106.
Haller, M., 1979. Estratigrafía de la región al poniente de Puerto Madryn, Provincia del Chubut, República Argentina. 7º Congreso Geológico Argentino, 1: 285-297. Neuquen, 1978.
Haller, M. y Mendía, J., 1980. Las sedimentitas del ciclo Patagoniano en el litoral atlántico nordpatagónico. En: Mendía, J.E. y Bayarsky, A., 1981. Estratigrafía del Terciario en el valle inferior del río Chubut. Actas del 8º Congreso Geológico Argentino, 3: 593-606.
Haq, B., Hardenbol, J. y Vail, P., 1988. Mesozoic and Cenozoic chronostratigraphy and cycles of sea-level change. En: Wilgus, Ch. y otros (Eds.) Sea level changes: An integrated approach. SEPM Sp. Publ., 42: 71-108.
Howard, J.D., 1966. Characteristic trace fossils in Upper Cretaceous sandstones of the Book Cliffs and Wasatch Plateau. Utha Geological and Mineralogical Survey, Bulletin 80: 35-53.
Howard, J.D. y Frey, R.W., 1975. Regional animal-sediment characteristics of Georgia estuaries. Senckenbergiana Maritima, 7: 33-103.
Howard, J.D. y Frey, R.W., 1984. Characteristic trace fossils in nearshore to offshore sequences, Upper Cretaceous of east-central Utah. Canadian Journal of Earth Sciences, 21: 200-219.
Ihering, H. von 1907. Les Mollusques fossiles du Tertiaire et du Cretacé Superieur de l´Argentina. Anales del Museo Nacional de Historia Natural. Serie 3 (7): 1-611.
Lech, R.R. y Buatois, L., 1990. Trazas fósiles del Pérmico marino de la Precordiller mendocina, República Argentina. 5º Congreso Argentino de Paleontología y Bioestratigrafía. Serie de Correlación Geológica, 7: 97-101.
Lesquereaux, L., 1876. Species of fossils marine plants from the Carboniferous measures. Indiana Geology Survey, Annual Report, 7: 134-145.
Lundgren, B., 1891. Studier öfver fossilförande lösa block. Geologiska Föreningens i Stockholm Förhandlingar, 13: 111-121.
Mángano, G. y Buatois, A., 1994. Trazas fósiles e icnofábricas en depósitos carbonáticos cretácicos, Las Cuevas, alta cordillera de Mendoza. Ameghiniana, 31 (1): 55-66.
Mendía, J.E. y Bayarsky, A., 1981. Estratigrafía del Terciario en el valle inferior del río Chubut. Actas del 8º Congreso Geológico Argentino, 3: 593-606.
Nicholson, H.A., 1873. Contributions to the study of the errant annelides of the older Palaeozoic rocks. Royal Society of London Proceedings, 21: 288-290.
Nicholson, H.A., 1873. Contributions to the study of the errant annelides of the older Palaeozoic rocks. Royal Society of London Proceedings, 21: 288-290.
d´Orbigny, A., 1842. Voyage dans l´Amérique méridionle (le Brésil, la République oriental de l´Uruguay, la République Argentine, la Patagonie, la République du Chili, la République de Bolivia, la République du Pérou) exécuté pendant les annees 1826, 1827, 1829, 1830, 1831, 1832, et 1833: v. 3, pt. 4 (Paléontologie), 188 p., 22 pl. Pitois - Levrault, París.
Ortman, A., 1902. Tertiary Invertebrate. Deports of the Princ. Univ. Expd. Patagonia, IV, 2.
Pemberton, S.G. y Frey, R.W., 1982. Trace fossil nomenclature and the Planolites - Palaeophycus dilemma. Journal of Paleontology, 56: 843-881. Tulsa.
Pemberton, S.G. y Jones, B., 1988. Ichnology of the Pleistocene Ironshore Formation, Gran Cayman Island, British West Indies. Journal of Paleontology, 62 (4): 495-505.
Pemberton, S.G., Frey, R.W. y Bromley, R.G., 1988. The ichnotaxonomy of Conostichus and other plug-shaped ichnofossils. Canadian Journal Earth Sciences, 25 (6): 866-892.
Pemberton, S.G., Van Wagoner, J.C. y Wach, G.D.; 1992. Ichnofacies of a wave-dominated shoreline. En Pemberton, S.G., 1992. Application of Ichnology to Petroleum Exploration. SEPM Core Workshop 117: 339-382.
Ranger, M.J. y Pemberton, S.G., 1988. Marine influence on the Mc Murray Formation in the Primrose area, Alberta. En: D.P. James y D.A. Leckie (Eds.). Sequences, Stratigraphy, Sedimentology: surface and sub-surface. Canadian society of Petroleum Geologist, 15: 439-450.
Rieth, A., 1932. Neue Funde spongeliomorpher Fucoiden aus dem Jura Schwabens. Geologische und Palaeontologische Abhandlungen, new series, 19: 257-294.
Rovereto, G., 1921. Studi di Geomorfologia Argentina: V, La penisola Valdéz. Bollettino della Societá Geologica Italiana, 40: 1-47.
Ruppert, E.E. y Barnes, R.D., 1996. Zoología de Invertebrados. Mc Graw-Hill Interamericana, sexta edición: 1114 p.
Scasso, R. y Castro, L.., 1999. Cenozoic phosphatic deposits in North Patagonia, Argentina: phosphogenesis, sequence stratigraphy and paleoceanography. Journal of South American Earth Sciences 12: 471-487.
Scasso, R. y Del Río, C., 1987. Ambiente de sedimentación, estratigrafía y proveniencia de la secuencia marina del Terciario superior de la región de península de Valdés. Revista de la Asociación Geológica Argentina, 42 (3-4): 291- 321. Buenos Aires.
Sheenan, P.M. y Schiefelbein, D.R.J., 1984. The tracefossil Thalassinoides from the Upper Ordovician of the eastern Great Basin: deep burrowing in the early Paleozoic. Journal of Paleontology, 58: 440-447.
Wilckens, O., 1905. Die Meeresablagerungen der Kreide und Tertiärformation in Patagonien. Neues Jahrbuch für Geologie und Paläontologie, 21.
Windhausen, A., 1919. Rasgos de la historia geológica de la planicie costanera en la Patagonia septentrional. Boletín de la Academia Nacional de Ciencias, 23: 319-364.
Windhausen, A., 1921. Informe sobre un viaje de reconocimiento geológico en la parte Nordeste del territorio del Chubut, con referencia especial a la cuestión de la provisión de agua de Puerto Madryn. Boletín del Ministerio de Agricultura de la Nación, 24, serie B, Geología: 9-72.
Recibido: 28 de mayo de 2000
Aceptado: 15 de noviembre de 2000