Análisis fitolítico de la Formación Paraná en la provincia de Entre Ríos
Alejandro F. ZUCOL 1 y Mariana BREA 1
Abstract.- PHYTOLITH ANALYSIS OF PARANÁ FORMATION IN ENTRE RÍOS PROVINCE. The phytolith analysis is used to reconstruct the paleoenvironment and paleobotanic composition during the deposition of the Paraná Formation in Entre Ríos province. A paleocomunity of palms is described for first time in this formation. The phytolith assemblage at the Puerto Alvear section was dominated by palm phytoliths, associated with Poaceae and Cyperaceae phytoliths; less frequent are the Podostemaceae phytoliths and sponge spicules. Comparison of phytolith morphology with those of extant taxa allowed recongnition an alluvial plain paleocomunity. The paleoclimatic conditions in the deposition of these sediment were tropical - subtropical and humid. A revision of the palm paleocomunity along the Cenozoic of this region is described, with special atention of its current distribution in the flora.
Key words: Phytoliths. Paraná Formation. Miocene. Arecaceae.
Palabras claves: Fitolitos. Formación Paraná. Mioceno. Arecaceae.
Introducción
Los estudios micropaleobotánicos de la Formación Paraná, hasta el momento, se han basado en su totalidad en análisis paleopalinológicos (Gamerro, 1981; Anzótegui y Garralla, 1982, 1986; Garralla, 1989). El presente, es un aporte al conocimiento paleobotánico de esta unidad mediante el estudio de sus asociaciones fitolíticas, y forma parte del análisis paleoecológico y paleoambiental de esta secuencia sedimentaria marina, que se está realizando en el sector comprendido en los departamentos Diamante y Paraná de la provincia de Entre Ríos.
A partir de los estudios paleopalinológicos previos, la paleoflora de la Formación Paraná se ha ido conociendo en forma de mosaico, como consecuencia del carácter local de estos estudios.
Con la finalidad de hallar una visión regional a estos conocimientos, los análisis fitolíticos que se están realizando se han planificado en dos transectas que abarcan la barranca del río Paraná en los departamentos de Diamante y Paraná respectivamente. Esta separación responde principalmente a las ya conocidas diferencias estratigráficas existentes en ambas áreas.
La palinoflora descripta se encuentra compuesta por elementos florísticos pertenecientes a diferentes unidades fisionómicas, los cuales pueden asociarse con integrantes de los actuales dominios fitogeográficos tropicales y subtropicales. Este esquema vegetacional describe tres tipos principales de comunidades: la comunidad hidrófila, representada por especies de las familias Azollaceae, Halograceae, Poaceae, Asteraceae, Polygonaceae, Onagraceae, Amaranthaceae; la comunidad higrófila, integrada por especies de las familias Polipodiaceae,
Araucariaceae y Podocarpaceae, Poaceae, Aquifoliaceae, Euphorbiaceae, Myrtaceae y Sapindaceae; y la comunidad xerófila compuesta por especies de las familias Poaceae, Asteraceae, Anacardiaceae y Mimosaceae.
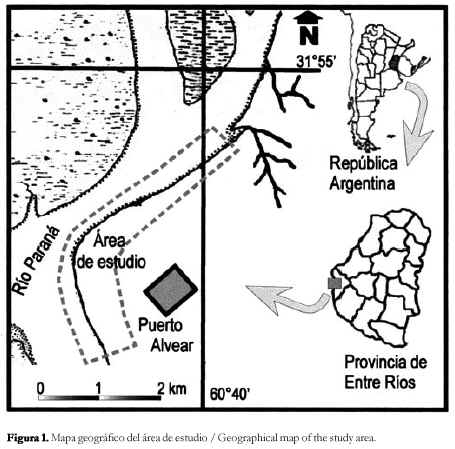
En este trabajo se aborda el estudio de los depósitos marinos en la región del Puerto General Alvear (departamento Diamante), ubicada a unos 30 km al S de la ciudad de Paraná sobre la margen izquierda del río homónimo (figura 1). Debe destacarse que esta localidad fosilífera ha permitido reconocer una nueva paleocomunidad vegetal que amplia los conocimientos paleoflorísticos de la Formación Paraná, incorporando nuevos registros a los ya descriptos.
La Formación Paraná en el perfil Puerto General Alvear
La Formación Paraná descripta por Bravard (1858), recibió diversas denominaciones nomenclaturales a lo largo del tiempo (Aceñolaza, 1976; 2000). Esta unidad de origen marino, se encuentra aflorante en las barrancas de Entre Ríos en una franja angosta que corre de N a S aproximadamente paralela al río Paraná. Litológicamente esta compuesta por arcillas, arcillas limosas y limos arenosos de color gris verdoso-oliva, con delgadas intercalaciones de arenas finas blanquecinas-amarillentas. Comúnmente se observan bancos calcáreos organógenos, constituidos principalmente por Ostrea patagonica. La asignación al Mioceno superior esta apoyada
por los estudios de Rossi de García (1966), Reinhart (1976), Zabert y Herbst (1977), Herbst y Zabert (1987) y Cozzuol (1993), sobre restos de vertebrados e invertebrados fósiles. Sin embargo, recientes estudios sobre las asociaciones de moluscos presentes en esta formación sugerirían que esta unidad fue depositada durante el Mioceno medio (Del Río, 1991); esta opinión es compartida por Aceñolaza y Aceñolaza (1999) basados en estudios de trazas fósiles. Aceñolaza (2000) describe y realiza la interpretación paleoambiental de esta unidad en detalle.
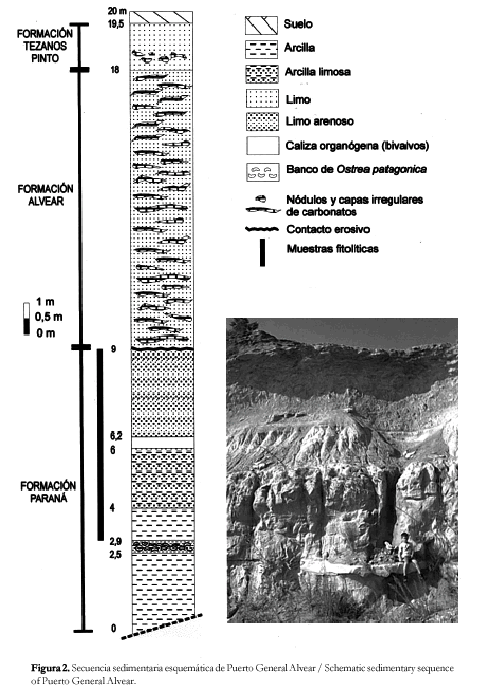
La secuencia sedimentaria (figura 2), en la localidad de Puerto General Alvear cuenta con un espesor de 20 metros, donde se observa de base a techo el siguiente perfil: La Formación Paraná, con un espesor de 9 m y sin base expuesta, se caracteriza por una importante sucesión de estratos arcillosos y areno-arcillosos de color gris verdoso-oliva, con delgadas intercalaciones de arenas finas blanquecinas-amarillentas, hacia el tope del perfil, estos estratos se vuelven más limosos y aumentan las intercalaciones arenosas, que a veces, se presentan en bancos masivos sin estratificación. Se observan los típicos bancos de ostreas y calizas organógenas portadoras de restos de bivalvos. Por encima se superpone en discordancia el calcrete típico de la Formacion Alvear (Pleistoceno inferior), constituido por nódulos y placas horizontales y verticales irregulares de carbonato de calcio de color blanquecino. Entre las placas, se encuentra un limo arcilloso pulverulento no calcáreo de color castaño rosado y castaño verdoso con nódulos y capas delgadas de manganeso. Hacia el techo, y con un espesor de 1,5 m se superponen los limos finos arcillosos pardos con nódulos de carbonato de calcio de la Formación Tezanos Pinto (Pleistoceno superior).
Análisis fitolítico
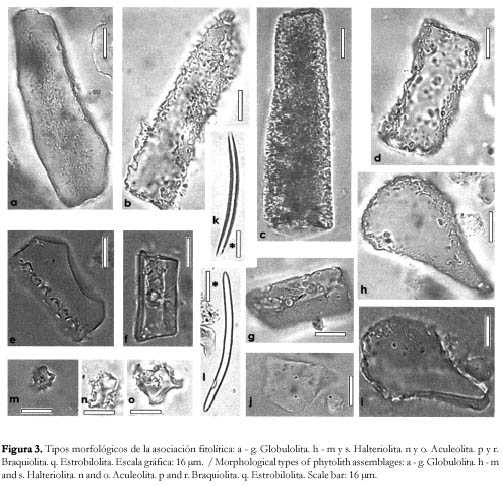
Asociación fitolítica. La asociación fitolítica se encuentra dominada por fitolitos esféricos (figura 3 a, b, d, y g), elipsoidales (figura 3 e y f) o ligeramente irregulares (figura 3 c), de diámetro variable entre 5 - 20 µm, con ornamentaciones cónicas cortas, pertenecientes a la morfotribu Globulolita (sensu Bertoldi de Pomar, 1971), asociados a fitolitos en forma de halterio (morfotribu Halteriolita) de extremos convexos (figura 3 h y j), rectos (figura 3 k y m), cóncavos (figura 3 i) o en forma de cruz de centro grueso (figura 3 l) o delgado (figura 3 s); fitolitos prismáticos, aplanados (morfotribu Prismatolita), con aristas laterales lisas (figura 4 a, c, d, f y g), denticuladas-serradas (figura 4 b) u ondulada (figura 4 e), y caras terminales superior e inferior planas, si bien se han observado algunos con caras cóncavas; fitolitos prismáticos aplanados con caras frontales flabeladas (morfotribu Flabelolita), de desarrollo simétrico (figura 4 i) o asimétrico (figura 4 h) respecto al plano longitudinal y fitolitos prismáticos cortos (morfotribu Braquiolita) con caras rectangulares de bordes lisos (figura 3 p y r). En menor frecuencia, se hallan presentes fitolitos en forma de aguijones o tricomas (morfotribu Aculeolita) de tamaño variable; fitolitos irregulares, escrobiculados (morfotribu Nuxolita) (figura 4 m, n y o); fitolitos pileiformes (morfotribu Pileolita) lisos u ornamentados; fitolitos en forma de conos o estróbilos (morfotribu Estrobilolita) truncados y laterlamente comprimidos (figura 3 q) y fitolitos de asignación incierta según la clasificación de Bertoldi de Pomar (figura 4 j).
Además de los fitolitos descriptos, se han hallado otros microrestos silíceos asociados entre los que son abundantes las espículas de espongiarios de agua dulce, principalmente del tipo de las macroescleras (tanto óxeas (figura 4 k y l) como anfiestróngilos), y en menor frecuencia, microescleras; por lo general aparecen enteras aunque a veces se observan macroescleras que-bradas.
En forma escasa, se observan también quistes de crisostomatáceas, de morfología afín al género Carnegia sp.
Afinidad botánica
Morfotribu Globulolita: si bien existen ciertos morfotipos de este grupo afines a las Poaceae (Globulolita claveteata, Bertoldi de Pomar, 1971), en su mayoría, y es ésta asociación el caso, son asignables a la familia Arecaceae.
Morfotribu Halteriolita: este grupo de fitolitos asignables en su totalidad a las Poaceae, en la asociación presentan un gran porcentaje de morfotipos de caracter panicoide (sensu Twiss, 1992), por sobre los pooides (sensu Twiss, 1992) y tipo stipa (sensu Fredlund y Tieszen, 1994).
Morfotribu Prismatolita: esta morfotribu en la asociación se encuentra mayoritariamente representada por fitolitos de afinidad graminoide y en bajo porcentaje de afinidad ciperoide.
Morfotribu Flabelolita: los fitolitos de esta morfotribu poseen afinidad graminoide y no permiten establecer diferenciaciones a nivel infrafamiliar.
Morfotribu Braquiolita: los fitolitos de esta morfotribu presentes en la asociación poseen exclusivamente afinidad graminoide.
Morfotribu Aculeolita: principalmente representado por fitolitos de afinidad graminoide, del tipo de los aguijones foliares marginales y escasamente en forma de ganchos.
Morfotribu Nuxolita: los fitolitos observados de esta morfotribu poseen en su totalidad afinidad con la familia Podostemaceae.
Morfotribu Pileolita: esta morfotribu se encuentra representada en la asociación por fitolitos de afinidad ciperoide.
Morfotribu Estrobilolita: los representantes fitolíticos de esta morfotribu poseen una afinidad graminoide, principamente del tipo chloridoide o más raramente pooide (sensu Twiss, 1992).
Paleoecología y paleoambientes
Mediante estos estudios se ha podido reconocer la presencia de nuevas familias para la Formación Paraná, tales como las Arecaceae y Podostemaceae, y corroborar otras descriptas en estudios paleopalinológicos. El análisis fitolítico permite definir una paleocomunidad dominada por palmeras asociadas a gramíneas (con predomino del tipo panicoide) y ciperáceas. Por otra parte la presencia de fitolitos de podostemáceas, indica la existencia de algunos cursos de aguas torrentosos, ya que en la actualidad estos vegetales se encuentran limitados a caídas, rápidos y cascadas de agua límpida (Bertoldi de Pomar, 1970). Por otra parte, la presencia de espículas de espongiarios de agua dulce con escaso grado de desintegración apoyan esta inferencia. El bajo porcentaje relativo de los fitolitos de ciperáceas y de lóricas de crisostomatáceas, están indicando que los cuerpos de agua lénticos eran escasos.
Los sedimentos estudiados son depósitos de una planicie aluvional, donde coexistían paleocomunidades terrestres como los palmares con paleocomunidades fluviales. Estas cond-
i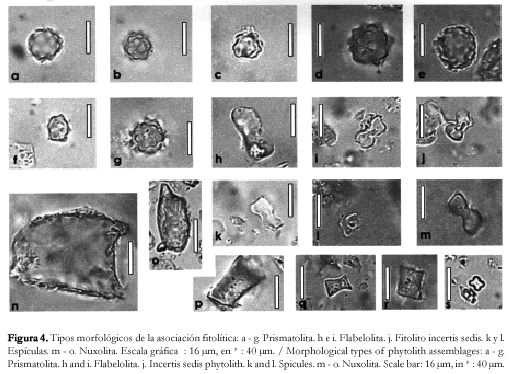
ciones alternaron con episodios cíclicos de incursiones marinas, como lo indica la presencia del banco de ostras infrayacente y la caliza organógena con restos de bivalvos. El palmar se desarrolló en un paleoambiente marino costero de llanura aluvional surcada por ríos o arroyos bastante correntosos con escasa presencia de ambientes inundables o anegados, y condiciones paleoclimáticas tropicales a subtropicales húmedas.
En correspondencia a estas inferencias, sobre la base de estudios litológicos, Iriondo (1973) concluye que esta secuencia sedimentaria fue depositada en un ambiente de poca profundidad muy cercano a la línea de costa correspondiendo a un depósito marino infralitoral a litoral.
Mientras que los estudios realizados sobre la icnofauna de esta unidad sugieren un ambiente marino litoral a sublitoral desarrollado bajo condiciones paleoambientales templadas a cálidas (Aceñolaza y Aceñolaza, 1999).
Esta paleocomunidad constituye el primer registro de este tipo para la Formación Paraná, y de acuerdo al esquema paleovegetacional descripto mediante análisis paleopalinológicos permite la incorporación de un ambiente de planicie aluvional (figura 5) donde se desarrolló una paleocomunidad dulceacuícola de cursos de agua correntosos que se encontraba presente junto con la paleocomunidad de los palmares.
Registros de la paleocomunidad de los palmares durante el Cenozoico superior y su distribución en la flora actual
Los primeros registros palinológicos de las Arecaceae datan del Maastrichtiano (Muller, 1979, 1981); en Sudamérica, se ha observado su presencia a lo largo de todo el Cenozoico (Menéndez, 1971; Romero, 1978 y 1986). En la región mesopotámica los registros más anti-

guos descriptos hasta el momento pertenecen a la Formación Ituzaingó (Plio-Pleistoceno) y Formación Salto Chico (Plio-Pleistoceno) con la descripción de granos de polen y leños petrificados, respectivamente (Anzótegui y Lutz, 1987). En la paleocomunidad descripta en este trabajo, se realiza la primera cita de esta familia para la Formación Paraná, ampliando su biocrón regional hasta el Mioceno medio.
En la secuencia sedimentaria del perfil Puerto General Alvear, a la Formación Paraná se superpone en discordancia la Formación Alvear (Pleistoceno inferior), en donde los estudios realizados muestran la presencia de fitolitos del tipo de las globulolitas. Estos fitolitos presentan una menor frecuencia relativa, y es notoria una mayor abundancia de fitolitos de afinidad graminoide, siendo estos últimos totalmente dominantes en la asociación fitolítica de la Formación Tezanos Pinto (Pleistoceno superior) suprayacente.
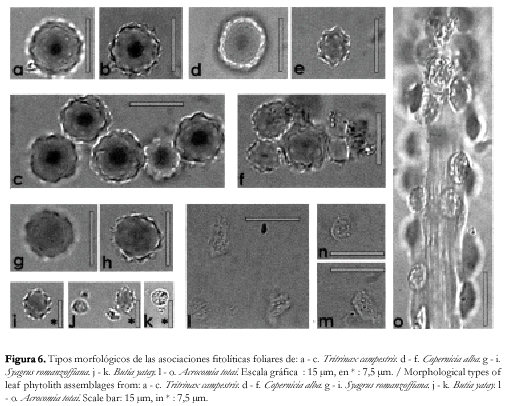
En lo que respecta a la distribución actual de los palmares, en la región se encuentran representadas dos tribus de las Arecaceae: las Corypheae (con las especies Trithrinax campestris y Copernicia alba) y las Cocoeae (con las especies Butia yatay y Syagrus romanzoffiana), de acuerdo a la clasificación de la familia propuesta por Uhl y Dransfield (1987) y Uhl et al. (1995). La distribución de estas especies ha sido bastante alterada durante el siglo pasado, tanto por la influencia antrópica como la climática; así relatos de la primera mitad de siglo (Báez, 1937 y 1942) describen a las especies de Trithrinax sp. y Copernicia sp. distribuidas en la región del Montiel, NE de Santa Fe (Ragonese, 1941) y Corrientes, y añade que "parece que la Caranday de Corrientes y Chaco, Copernicia y otras Trithrinax avanzaran por las llanuras chaqueñas hasta llegar al altiplano de Bolivia y quizá a los valles de la precordillera andina" (Báez, 1937: 70); en tanto Butia sp. y Syagrus sp. poseen una dispersión mesopotámica, con una abundancia que disminuye notoriamente hacia el S, formando comunidades restringidas que Báez denominó núcleos de palmares.
Al respecto, tanto Báez (op cit.) como Ibáñez (1962), señalan la presencia de un núcleo de B. yatay en la localidad de Puerto General Alvear, el cual es inexistente en la actualidad.
Sus características mesológicas muestran que T. campestris habita en las zonas de suelos arcillosos de la provincia de Entre Ríos, en climas templados cálidos, lluviosos, aunque se adapta a períodos de sequía, asociada a la vegetación de monte del NO de la provincia. C. alba se distribuye en lugares bajos y generalmente salobres de la provincia de Santa Fe (Ragonese, 1941). B. yatay es una palmera que crece en suelos muy arenosos; exceptuando los núcleos de la costa del río Uruguay, la especie no se distribuye en comunidades puras, con escasa importancia numérica, desarrollándose principalmente en albardones arenosos asociadas con matorrales arbustivos (Ibáñez, 1962). S. romanzoffiana es una palmera hidrófila, sociable con otros árboles ribereños, que crece preferentemente en terrenos húmedos y arenosos. Su origen es austrobrasileño, y su vía de penetración a la provincia de Entre Ríos es a través de la cuenca del río Uruguay (Báez, 1937; Ibáñez, 1962). De lo expuesto puede concluirse que las Arecaceae en la flora entrerriana están representadas por dos grupos: el de la tribu Corypheae con características afines al Dominio Chaqueño y el de la tribu Cocoeae con características de afinidad con el Dominio Amazónico.
El análisis comparativo de sus asociaciones fitolíticas permitió establecer que T. campestris (figura 6 a, b y c) se caracteriza por la presencia de fitolitos globulares espinosos de 14 - 20 µm de diámetro promedio. La asociación fitolítica de C. alba (figura 6 d, e y f) posee elevada abundancia de fitolitos globulares espinosos y microespinosos de elevado diámetro (14 - 20 µm) con menor frecuencia de fitolitos globulares espinosos de menor tamaño (7 - 10 µm de diámetro). Por otra parte, S. romanzoffiana presenta una asociación dominada por fitolitos globulares espinosos grandes (12 µm de diámetro promedio) (figura 6 g y h) y pequeños (7µm de diámetro promedio) (figura 6 i). B. yatay se caracteriza fitolíticamente por la presencia de cuerpos elípticos espinosos (figura 6 j) asociados con cuerpos circulares achatados en sus bordes (figura 6 k), morfológicamente similares a los observados en Acrocomia totai (figura 6 l, m, n y o).
De acuerdo al marco morfológico observado para las Arecaceae de la región, los fitolitos que componen la asociación descripta para la Formación Paraná, presentan una mayor afinidad con los representantes de la tribu Corypheae, cuyos integrantes son característicos del Dominio Chaqueño.
Agradecimientos
Los autores desean agradecer al Dr. R. Herbst cuya lectura crítica y sugerencias permitieron mejorar el manuscrito . Este trabajo se ha realizado con el financiamiento del PEI-CONICET 0113-98.
Bibliografía
Aceñolaza, F.G. 1976. Consideraciones bioestratigráficas sobre el Terciario marino de Paraná y alrededores. Acta Geológica Lilloana 13 (2): 91-107. Tucumán.
Aceñolaza, F.G. 2000. La Formación Paraná (Mioceno medio): estratigrafía, distribución regional y unidades equivalentes. En: Aceñolaza F. G. y Herbst, R. (eds.), El Neógeno de Argentina. Correlación Geológica 14: 9-27.
Aceñolaza, F.G. y Aceñolaza, G.F. 1999. Trazas fósiles del Terciario marino de Entre Ríos (Formación Paraná, Mioceno medio), República Argentina. Boletín de la Academia Nacional de Ciencias , Cordóba 64: 209-233. Cordóba.
Anzótegui, L.M. y Garralla, S.S. 1982. Estudio palinológico de la Formación Paraná (Mioceno superior). I Parte. Pozo "Josefina", provincia de Santa Fe, Argentina. III Congreso Argentino de Paleontología y Bioestratigrafía, Resumen. : 32. Corrientes.
Anzótegui, L.M. y Garralla, S.S. 1986. Estudio palinológico de la Formación Paraná (Mioceno superior) (Pozo "Josefina", provincia de Santa Fe, Argentina. I Parte - Descripción sistemática. Facena, 6: 101-177.
Anzótegui, L.M. y A.I. Lutz. 1987. Paleocomunidades vegetales del Terciario superior ("Formación Ituzaingó") de la Mesopotamia Argentina. Revista de la Asociación de Ciencias Naturales del Litoral, 18(2): 131-144.
Báez, J.R. 1937. Área de dispersión actual de las palmeras en la flora de Entre Ríos. Anales de la Sociedad Argentina de Estudios Geográficos "GEA", 5: 63-78.
Báez, J.R. 1942. Las palmeras en la flora de Entre Ríos. Public. Comis. Coordinadora de Servicios. Paraná. Exposic. forestal de Bs. As (M. A.N.), :1-11.
Bertoldi de Pomar, H. 1970. F itolitos y zoolitos, su significado geológico en sedimentos continentales. Boletín de la Asociación Geológica de Córdoba, 1(1): 21-31.
Bertoldi de Pomar, H. 1971. Ensayo de clasificación morfológica de los silicofitolitos. Ameghiniana, 8 (3-4): 317-328.
Bravard, A. 1858. Monografía de los terrenos marinos terciarios de las cercanías del Paraná. Imprenta del Registro Oficial. 107 pp. Paraná. (Reimpresión Imprenta del Congreso de la Nación, 1995).
Cozzuol, M.A. 1993. Mamíferos acuáticos del Mioceno medio y tardío de Argentina. Sistemática, evolución y biogeografía. Tesis Doctoral, Facultad de Ciencias Naturales y Museo, UNLP. 178 pp. Inédita.
Del Río, C. 1991. Revisión sistemática de los bivalvos de la Formación Paraná (Mioceno medio) provincia de Entre Ríos. Monografía de la Academia Nacional de Ciencias Exactas, Físicas y naturales, 7, 93 pp. Buenos Aires.
Fredlund G. y L.T. Tieszen. 1994. Modern phytolith assemblages from the North American Great Plains. Journal of Biogeography, 21: 321-335.
Gamerro, J.C. 1981. Azolla y Salvinia (Pteridiphyta Salviniales) en la Formación Paraná (Mioceno superior), Sta Fe, Rep. Argentina. IV Simposio Argentino de Paleobotánica y Palinología, Resumen, 3: 12-13.
Garralla, S.S. 1989. Palinomorfos (Fungi) de la Formación Paraná (Mioceno superior) del pozo Josefina, provincia de Santa Fe, Argentina. Revista de la Asociación de Ciencias Naturales del Litoral, 20(1-2) :29-39.
Herbst, R. y Zabert, L.L. 1987. Microfauna de la Formación Paraná (Mioceno superior) de la cuenca Chaco-Paranense (Argentina). Facena 7: 165-206. Corrientes.
Ibáñez, F.M. 1962. Vegetación de la provincia de Entre Ríos. Revista Nordeste, 4 : 93-127.
Iriondo, M. 1973. Análisis ambiental de la Formación Paraná en su área tipo. Boletín de la Asociación Geológica de Córdoba, 2 (1): 19-24.
Menéndez, C.A. 1971. Floras terciarias de la Argentina. Ameghiniana, 8(3-4): 357-371.
Muller, J. 1979. Reflections of fossil palm pollen. En: 4 th International Palynological Conference: proceedings, 1: 568-578. Birbal Sahni Institute of Paleobotany, Lucknow.
Muller, J.1981. Fossil pollen records of extant angiosperms. Botanical Review, 47:1-142.
Ragonese, A.E. 1941. La vegetación de la provincia de Santa Fe. Darwiniana 5: 369-428.
Reinhart, R. 1976. Fossil Sirenians and Desmostylides from Florida and Elsewhere. Bulletin Florida State Museum of Biological Science, 20: 187-300. Florida.
Romero, E. 1978. Paleoecología y paleofitogeografía de las tafofloras del Cenofítico de Argentina y áreas vecinas. Ameghiniana, 15(1-2): 209-227.
Romero, E. 1986. Paleogene phytogeography and climatology of South America. Annals of the Missouri Botanical Garden, 73: 449-461.
Rossi de García, E. 1966. Contribución al conocimiento de los ostrácodos de la Argentina. I. Formación Entre Ríos de Victoria, provincia de Entre Ríos. Asociación Geológica Argentina, Revista 21 (3): 194-208. Buenos Aires.
Twiss, P.C. 1992. Predicted world distribution of C 3 and C 4 grass phytoliths. En: Rapp, G. and S.C. Mulholland (eds.): "Phytolith Systematics." :113-128. Plenum Press.
Uhl, N.W. y J. Dransfield. 1987. Genera palmarum: a clasification of palms based on the work of H.E. Moore, Jr. 610 pp. International Palm Society and L.H. Bailey Hortorium, Lawrence, Kan. Uhl, N.W.; Dransffield, J.; Davis, J.I.; Luckow, M. A.; Hansen, K.S. y J.J. Doyle. 1995. Phylogenetic relationships among palms: Cladistic analyses of morphological and chloroplast DNA restriction site variation. En: Rudall, P.J.; Cribb, P.J.; Culter, D.F. y C.J. Humphries (eds.): "Monocotyledons: systematics and evolution". :623-661. Royal Botanical Garden, Kew. Zabert, L.L y Herbst, R. 1977. Revisión de la microfauna Miocena de la Formación Paraná (entre Victoria y Villa Urquiza, provincia de Entre Ríos, Argentina) con algunas consideraciones estratigráficas. Facena 1: 131-164. Corrientes.
Recibido: 18 de agosto de 2000
Aceptado: 20 de noviembre 2000