 ambientes
marinos muy someros, normales a hiposalinos, y de una temperatura similar o algo
superior a la actual para los mares
adyacentes. Los foraminíferos pertenecen a la zona informal o
hemerozona de Protelphidium tuberculatum (Malumián,
1970).
ambientes
marinos muy someros, normales a hiposalinos, y de una temperatura similar o algo
superior a la actual para los mares
adyacentes. Los foraminíferos pertenecen a la zona informal o
hemerozona de Protelphidium tuberculatum (Malumián,
1970).Rasgos micropaleontológicos de los depósitos de la transgresión Entrerriense-Paranense en la cuenca Chaco-Paranense y Noroeste Argentino.
Hugo Guillermo MARENGO 1
Abstract.- MICROPALEONTOLOGICAL OUTLINES OF THE ENTRERRIENSE-PARANENSE TRANSGRESSION BEDS IN THE CHACO-PARANENSE BASIN AND THE NORTHWEST ARGENTINA. From the Middle Miocene to the Upper Miocene an extended transgression flooded a significant portion of the Argentinian territory, and it reached the south of Bolivia and Paraguay. This transgression has been generally known as the entrerriense or paranense transgression (TEP). This study summarizes all the available information about calcareous marine microfossils deposited by the TEP in the NW argentine region, Chaco-Paranense basin, and west Mesopotamia, Argentina. 66 cores recovered from four drillings on the Diamante area, Entre Ríos province, have been analyzed, and they constitute the first subsurface study, and the first study of the complete sequence of the Paraná Fm in its type area.
Foraminiferal assemblages belong of the Protelphidium tuberculatum informal Zone. P. tuberculatum and Ammonia parkinsoniana are widely dominant in the studied areas. In almost all regions the microfaunas are typical of shallow and brackish waters, but in some localities of SW Entre Ríos province and central Santa Fe province, microfaunas that indicate normal marine conditions have been recognized. TEP foraminiferal assemblages are very similar to those wich live today in adjacent seas, with scattered specimens of warmer water species; consequently, the water temperature should have been the same, or just a few degrees higher than at the today adjacent seas. In the Diamante area, marine sedimentation was interrupted, in the lower and middle Paraná Formation, by prograding coastal systems (probably deltaics). To the top of the section, and taking account the richness and abundance of microfossils, the sea level had reached its higher level.
The foraminiferal and ostracod geographical distribution indicate that the sea entered from the Atlantic through the Salado basin. The hypothesis here proposed argue against the faunal migration through an hypothetic intracontinental seaway connecting the Caribbean sea with the Plata region during TEP deposition, known as the "Arm of Tethys".
Key words: Chaco-Paranense Basin, Middle-Late Miocene, Marine Calcareous Microfossils, Paleogeography, Paleoenvironments, Eustatic changes.
Palabras clave: Cuenca Chaco-Paranense, Mioceno medio-tardío, microfósiles calcáreos marinos, paleogeografía, paleoambientes, variaciones eustáticas.
Introducción
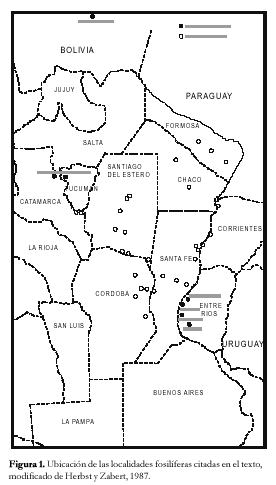
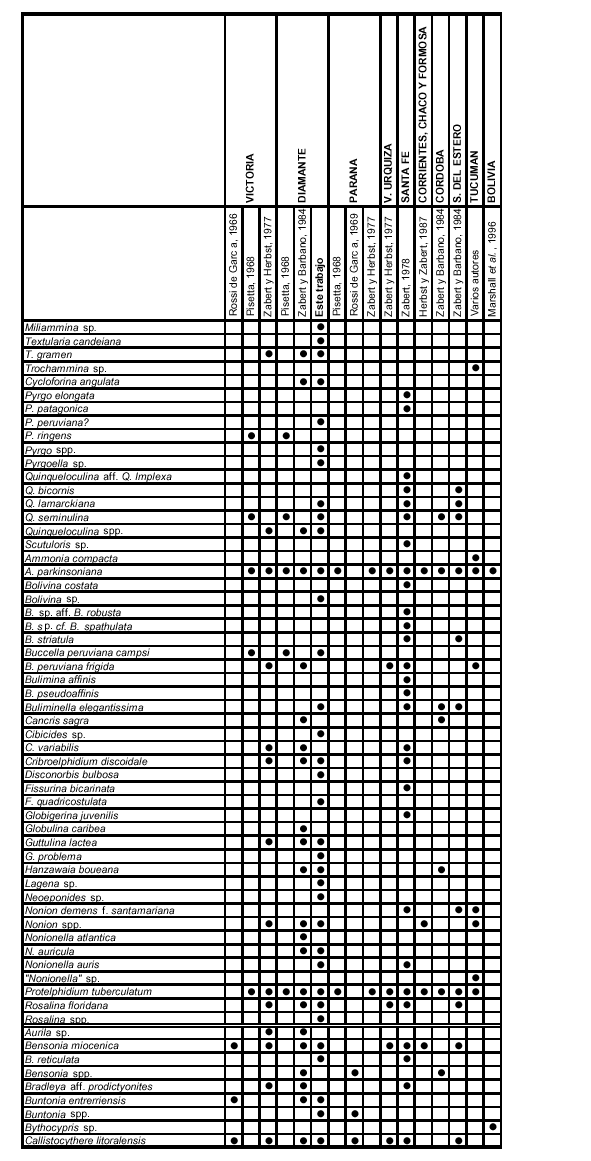
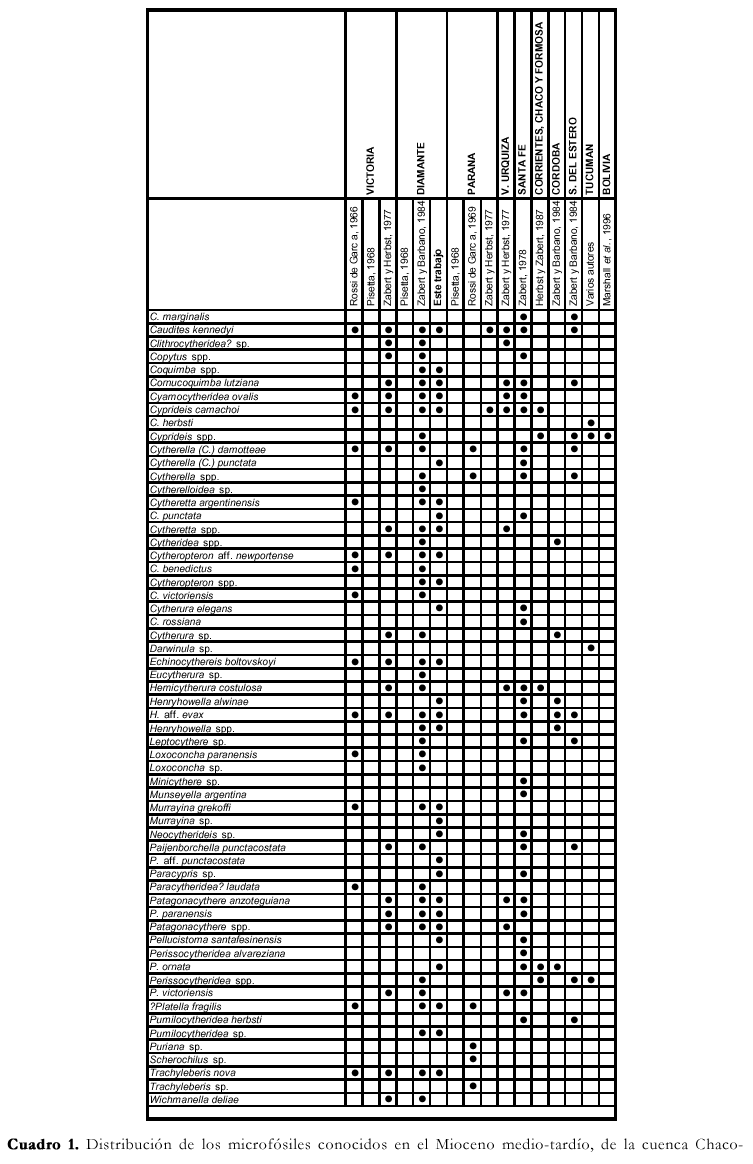
Durante el Mioceno medio-tardío ?, una transgresión cubrió gran parte del territorio argentino, parte de las costas del Uruguay y del sur del Brasil, y el sur de Bolivia y Paraguay (Figura 3), reconocida en distintas Formaciones, según los autores y las regiones (Fm. Paraná en cuenca Chaco-Paranense, «arcillas verdes» en la cuenca del Salado, Fm. Barranca Final en la cuenca del Colorado, Fm. Puerto Madryn en el NE del Chubut, Fm. Macachín en la cuenca homónima, Fm San José en las provincias de Tucumán y Catamarca, Fm. Yecua en el sur de Bolivia, Fm. Camacho en el Uruguay, etc.; además es conocida con los nombres genéricos de «entrerriense» o «paranense»). En este trabajo se la denominará, en sentido amplio, Transgresión Entrerriense-Paranense (TEP). El contenido micropaleontológico de la TEP ha sido estudiado en casi todas las regiones: subsuelo de península de Valdés (Masiuk et al., 1976), afloramientos del NE de la provincia de Río Negro (Malumián et al., 1998), cuenca del Colorado (Malumián, 1970 y 1972; Becker y Bertels, 1980; Boltovskoy, 1980), cuenca del Salado (Malumián, 1970 y 1972), afloramientos del SO de la provincia de Entre Ríos (Rossi de García, 1966 y 1969; Pisetta, 1968; Zabert y Herbst, 1977; Zabert y Barbano, 1984), cuenca Chaco-Paranense (Zabert, 1978; Zabert y Barbano, 1984; Herbst y Zabert, 1987), afloramientos de las provincias de Tucumán y Catamarca (Bertels y Zabert, 1980; Gavriloff et al., 1998), y del sur de Bolivia (resumen en Marshall et al., 1993).
Se resume toda la información disponible sobre los foraminíferos y ostrácodos de la cuenca Chaco-Paranense, Noroeste Argentino y sur de Bolivia, y se analiza su significado paleoambiental y su distribución geográfica.
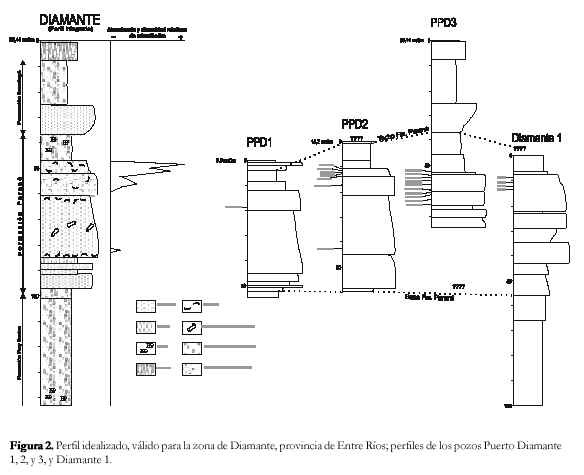
Si bien existen varios estudios sobre las microfaunas del SO de Entre Ríos, y en consecuencia el conocimiento de su composición taxonómica es extenso, corresponden en todos los casos a perfiles de afloramientos en los que no afloran las secciones inferiores ni los contactos basales de la Formación Paraná. Para subsanar esta falta, se estudiaron los microfósiles de los testigos de cuatro perforaciones de la zona de Diamante.
Sobre la base de la comparación de todas las microfaunas conocidas en la región se intenta establecer la dinámica de las poblaciones de microfósiles, su distribución geográfica y estratigráfica, y las características generales de la transgresión.
Materiales y métodos
En el cuadro de distribución general de la cuenca Chaco-Paranense, y en el apéndice, se ha mantenido la sinonimia original con la menor cantidad posible de variantes; sólo aquellos taxones que a criterio del autor son idénticos se unificaron bajo un único nombre. Los géneros de los foraminíferos han sido actualizados según Loeblich y Tappan (1988); para los ostrácodos se mantienen los géneros dados por los autores que los reconocieron previamente en la región.
Los foraminíferos se determinaron mediante la comparación con material de la colección de micropaleontología del SEGEMAR y con la bibliografía específica. La determinación de los ostrácodos es preliminar, debido a que sólo se contó con material bibliográfico.
Se seleccionaron los testigos de las perforaciones más profundas, y con una mayor densidad en la toma de las muestras. De los numerosos testigos de perforación de la provincia de Entre Ríos depositados en el repositorio del SEGEMAR, los de la zona de Diamante resultaron los más apropiados. El muestreo fue realizado por el método de la cuchara; debido a que los intervalos de la toma de las muestras son muy irregulares, se estudiaron varias perforaciones, con el fin de obtener una sección promedio representativa de la zona de Diamante.
No se conoce la ubicación geográfica exacta de las perforaciones; de Puerto Diamante 1, 2, y 3 se conocen las cotas, medidas en metros sobre el cero del mareógrafo del Riachuelo; de la perforación Diamante 1 no se conoce dato alguno. Se asume que todas las perforaciones fueron realizadas lo suficientemente cercanas entre sí, de modo que no poseen diferencias estratigráficas importantes. Las mayores diferencias entre los perfiles se atribuyen a la cota a que fueron realizadas las perforaciones, con la lógica variación en el espesor de la cubierta cuaternaria reciente.
La información obtenida de la descripción de los testigos de perforación, se complementó con los perfiles de superficie de Pisetta (1968) y Zabert y Herbst (1977).
El procesamiento de las muestras fue el habitual en los estudios micropaleontológicos: desagregado con agua oxigenada 20 vol. durante 24 hs., lavado en tamiz de 62,5 micrones, separación de todos los ejemplares de microfósiles de la fracción mayor a 62,5 micrones bajo lupa binocular. Se estudiaron muestras de peso estándar, con el objetivo de facilitar la comparación entre las diferentes asociaciones.
Antecedentes
Todos los trabajos previos sobre la micropaleontología de la Fm. Paraná y equivalentes poseen características similares, ya que solamente se realizaron estudios sistemáticos, y no se conocen la abundancia relativa ni la variación vertical de los diferentes taxones (Cuadro 1).
En
los afloramientos del SO de la provincia de Entre Ríos, Rossi de García (1966
y 1969), Pisetta (1968), Zabert y Herbst
(1977), y Zabert y Barbano (1978), hallaron microfaunas de ostrácodos
y foraminíferos caracterizadas por su baja
a moderada diversidad, indicadoras de ambientes
marinos muy someros, normales a hiposalinos, y de una temperatura similar o algo
superior a la actual para los mares
adyacentes. Los foraminíferos pertenecen a la zona informal o
hemerozona de Protelphidium tuberculatum (Malumián,
1970).
Zabert (1978) y Herbst y Zabert (1987) estudiaron las microfaunas de la Formacion Paraná en el subsuelo de la provincia de Santa Fe. Sus resultados paleoambientales y paleoecológicos coinciden plenamente con los dados para el SO de Entre Ríos, aunque la composición taxonómica indicaría una profundidad del mar levemente mayor y una salinidad más cercana a la normal, principalmente por la aparición de los géneros Globigerina, Bolivina y Bulimina, y por una mayor diversidad de foraminíferos miliólidos.
Las microfaunas del subsuelo de las provincias de Córdoba y Santiago del Estero (Zabert y Barbano, 1984; Herbst y Zabert, 1987), de Corrientes, Chaco, y Formosa (Herbst y Zabert, 1987), y de afloramientos del valle de Santa María, provincia de Tucumán (Bertels y Zabert, 1980; Gavriloff et al., 1998) están marcadamente empobrecidas en diversidad, y son características de aguas muy hiposalinas, con un dominio casi exclusivo de P. tuberculatum y Ammonia parkinsoniana.
Marshall et al. (1993) resumen los hallazgos micropaleontológicos, inéditos, en el sur de Bolivia. Estos se restringen solamente a una especie de foraminífero bentónico de amplia tolerancia a los cambios de salinidad, A. parkinsoniana, y a dos géneros de ostrácodos de agua dulce.
Por último, Malumián (1970 y 1972) estudió los foraminíferos del "entrerriense" del subsuelo de la cuenca del Salado, depósitos que si bien no se incluyen en la presente contribución, están íntimamente ligados a los del sur de la cuenca Chaco-Paranense. Este autor describió microfaunas muy similares a las del sur de las provincias de Santa Fe y Entre Ríos, aunque más diversas y con una mayor cantidad de foraminíferos planctónicos, y corresponderían a un mar menos restringido y más profundo. Agrupó estos foraminíferos en la zona informal o hemerozona de P. tuberculatum, caracterizada por foraminíferos bentónicos de mares someros, con una muy baja proporción de miliólidos y aglutinados, y dominada por P. tuberculatum, Cribroelphidium discoidale, Buccella peruviana campsi y Hanzawaia boueana.
Sobre la base de los antecedentes citados se puede resumir que: - La TEP fue un mar somero, con una marcada disminución de la salinidad y la profundidad, desde la cuenca del Salado hacia el noroeste y noreste Argentino, y Bolivia; la temperatura del mar fue similar o levemente superior a la de los mares adyacentes en la actualidad. La fauna de la zona informal de P. tuberculatum, característica de la cuenca del Salado y del sur de la cuenca Chaco-paranense, se empobrece rápidamente hacia el interior del continente, reduciéndose a muy pocas especies, generalmente tolerantes a las bajas salinidades, entre las que dominan ampliamente A. parkinsoniana y P. tuberculatum.
- La ausencia de fósiles guía, no ha permitido precisar la edad de la TEP en la región considerada.
Los diversos autores la han atribuido, en forma amplia, al Mioceno, más probablemente al Mioceno medio-tardío. La datación más confiable hasta la actualidad es la dada por Malumián (1970 y 1972), que sobre la base del hallazgo de Cassigerinella chipolensis (Cushman y Ponton) y Globigerina woodi woodi Jenkins, ubicó al "entrerriense" de la cuenca del Colorado, en el Mioceno medio, aunque considera que los depósitos de la cuenca del Salado serían del Mioceno tardío.
- El conocimiento del contenido micropaleontológico de la TEP en la cuenca Chaco-paranense y el noroeste Argentino es bastante amplio en cuanto a la composición taxonómica, pero en ningún caso se han proporcionado cuadros de distribución completos ni los valores numéricos de los ejemplares hallados.
Microfósiles del subsuelo de Diamante
Como se expuso anteriormente, los estudios micropaleontológicos del SO de Entre Ríos son abundantes, aunque todos fueron realizados en secciones en las que sólo afloran unos pocos metros; por lo tanto, no se conoce el contenido micropaleontológico ni la potencia de la sección inferior.
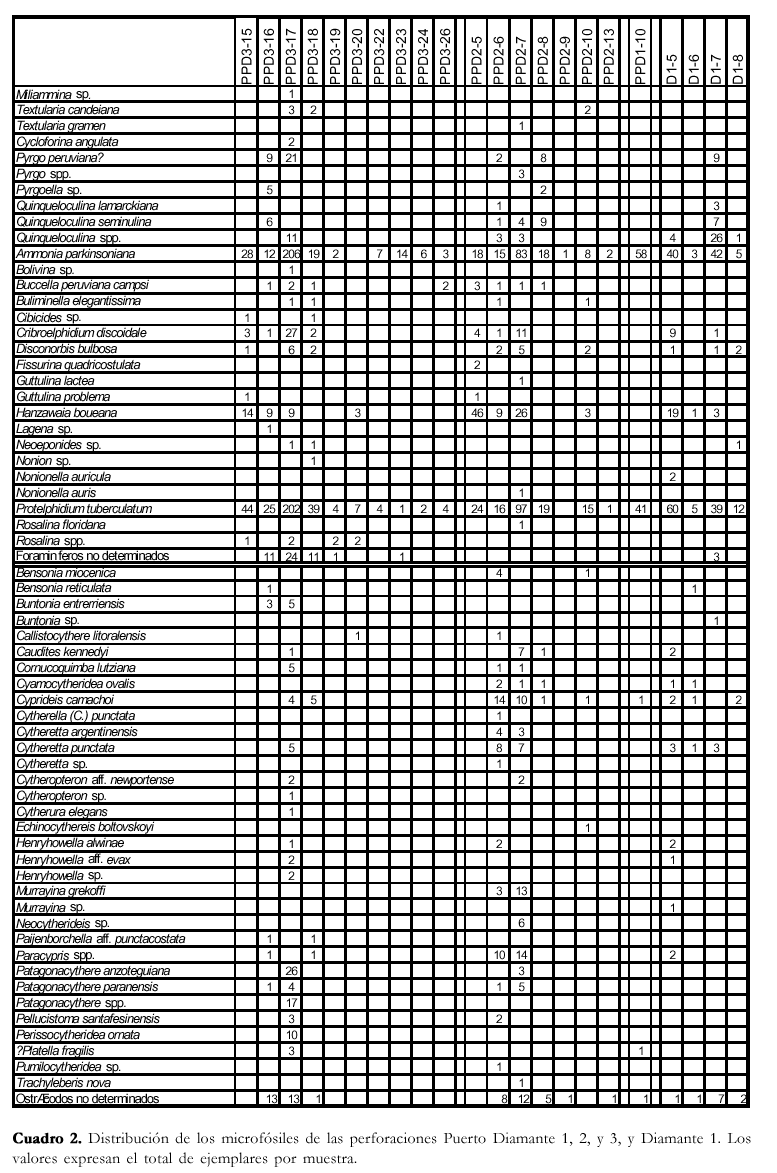
Se han seleccionado los testigos de las perforaciones Puerto Diamante 1, 2, y, 3, y Diamante 1, realizadas por la Dirección Nacional de Geología y Minería en la localidad de Diamante, provincia de Entre Ríos. Debido a la notable falta de uniformidad en la toma de las muestras, y a la diferente posición estratigráfica de las perforaciones, fue necesario realizar un perfil integrado, válido para la zona de Diamante (Figura 2), en donde se complementó la información litológica y micropaleontológica de subsuelo, con las descripciones de los perfiles de los aflora-mientos cercanos, realizadas por Pisetta (1968) y Zabert y Herbst (1977).
A continuación se describe el perfil integrado de la región de Diamante; las profundidades están expresadas en metros bajo boca de pozo, ubicada a 56,44 metros sobre el cero del mareógrafo del Riachuelo. 0,0-0,7 mbbp: Suelo castaño oscuro. Estéril.
0,7-7,5 mbbp: Limo masivo castaño claro, con fragmentos de tosca de unos 5 mm. Estéril.
7,5-25,0 mbbp: Pelita masiva castaña rojiza clara, con pequeñas concreciones calcáreas, posiblemente de tosca. Estéril.
25,0-37,0 mbbp: Arenisca masiva gris amarillenta de grano medio, fuertemente cementada por carbonato de calcio. Estéril.
37,0-47,0 mbbp: Pelita verde oliva clara, ocasionalmente laminada, con concreciones calcáreas y cristales de yeso. Estéril.
47,0-52,0 mbbp: Arenisca gris amarillenta de grano muy fino y poco seleccionada, muy friable.
Numerosos fragmentos de pelecípodos, y algunos ejemplares enteros de pequeñas ostras, de hasta 3 cm. Briozoarios, foraminíferos y ostrácodos muy abundantes; generalmente bien conservados, excepto en el caso de los foraminíferos miliólidos, parcialmente disueltos.
52,0-61,0 mbbp: Arenisca amarilla de grano medio y bien seleccionada, muy friable. Algunos fragmentos de ostras y otros moluscos indeterminables. Foraminíferos y ostrácodos escasos, generalmente recristalizados en diferentes grados.
61,0-85,0 mbbp: Arenisca amarilla a verde amarillenta de grano fino a grueso, muy friable, con delgadas intercalaciones de pelitas de tonalidades oscuras. Fragmentos de ostras muy recristalizadas en la base y el techo. Muy escasos foraminíferos y ostrácodos, muy recristalizados.
En algunas muestras se hallaron fragmentos de madera muy poco mineralizada, de hasta 2 cm.
85,0-97,5 mbbp: Intercalaciones de arenisca limosa gris verdosa con pelita arenosa verde oliva.
Estéril.
97,5-101,0 mbbp: Pelita castaña verdosa oscura con concreciones calcáreas. Estéril.
101,0-143,0 mbbp: Pelita masiva castaña rojiza oscura con concreciones calcáreas, y cristales de yeso en la base. Estéril.
Los microfósiles se distribuyen en forma similar en las cuatro perforaciones (Cuadro 2), por lo tanto se puede considerar, en líneas generales, la siguiente distribución para la zona de Diamante: ~ 47-53 mbbp: microfaunas de diversidad moderada, dominadas por P. tuberculatum, A. parkinsoniana, H. boueana, foraminíferos miliólidos, Paracypris sp., Cyprideis camachoi, Cyamocytheridea ovalis y Patagonacythere spp. Hacia el techo del banco disminuye la diversidad, con un considerable aumento de la proporción de H. boueana, y la desaparición de los miliólidos y los ostrácodos.
~ 55-60 mbbp: microfaunas de diversidad y abundancia muy bajas, dominadas por P. tuberculatum y A. parkinsoniana, y con unos pocos ejemplares de H. boueana, B. peruviana campsi, C. discoidale, y ostrácodos.
~ 85 mbbp: muy escasos ejemplares de P. tuberculatum y A. parkinsoniana.
Discusión
i) Características generales de las microfaunas. Las características generales de las microfaunas fueron puntualizadas por varios autores, y sintetizadas, dentro de un esquema regional, por Herbst y Zabert (1987). La información obtenida durante el presente trabajo no permite rectificar ni ampliar demasiado las conclusiones paleoambientales de los trabajos previos.
En resumen, las microfaunas de la Fm. Paraná, y equivalentes, son típicas de mares muy
someros, generalmente hiposalinos, y de una temperatura similar a algo superior a la actual para los mares adyacentes. Se hallaron algunos foraminíferos nuevos para la región, como Miliammina sp., Textularia candeiana, Pyrgoella sp., Disconorbis bulbosa, Fissurina quadricostulata, Guttulina problema, Lagena sp. y Neoeponides sp., pero no permitieron obtener conclusiones paleoambientales más precisas, excepto D. bulbosa, foraminífero bentónico actualmente restringido a las costas del norte del Brasil, que confirmaría que la temperatura de las aguas de la TEP era levemente mayor a la actual.
Los foraminíferos dominantes en todas las zonas consideradas son P. tuberculatum y A. parkinsoniana, especies de una amplia tolerancia a las bajas salinidades (Malumián, 1978; Murray, 1991). En el sector superior de la Fm. Paraná, en la zona de Diamante, se registraron microfaunas típicas de aguas de salinidad cercana a la normal, donde la diversidad de los ostrácodos y
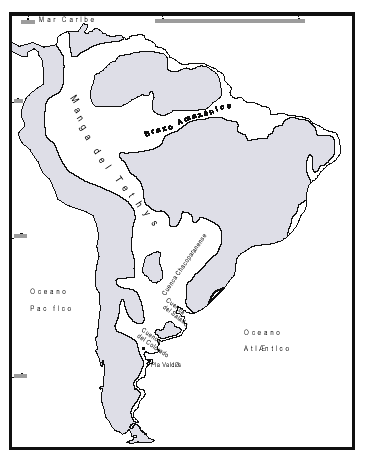
Figura 3. Paleogeografía de América del Sur Durante el Mioceno medio-tardío. Se incluye el hipotético brazo marino conocido como «Manga del Tethys». Modificado de Windhausen (1931), Uliana y Biddle (1988), y Pérez y Ramos (1996).
foraminíferos es máxima, y hay un notable incremento en la abundancia y diversidad de los miliólidos; en consecuencia, en este sector, la TEP habría alcanzado su máxima profundidad y el período de mayor inundación.
Las microfaunas con mayor diversidad se reconocieron en el SO de Entre Ríos y en el este de Santa Fe; hacia el NO y NE las microfaunas se empobrecen en forma notable, debido a la disminución de la profundidad del mar, y a la mezcla del agua marina con aguas dulces.
ii) La TEP en Diamante. La TEP habría comenzado, en la región de Diamante, como un lento ascenso del mar, del que sólo se registra un delgado nivel con fragmentos de ostras y muy escasos microfósiles.
Sobre este, se depositó un potente nivel de arenas gruesas a medianas, con pelitas intercaladas, características de un ambiente de transición. Frenguelli (1920), explica las intercalaciones de niveles arenosos en la Formación Paraná como producto de varios ciclos transgresivo-regresivos, mientras que Stappenbeck (1926) interpreta que fueron producidos por la progradación de un sistema deltaico sobre el ambiente costero; el hallazgo de fragmentos de madera y de intercalaciones de delgados bancos de pelitas oscuras apoyaría la interpretación de Stappenbeck (op. cit.) Aceñolaza y Aceñolaza (1999), por otro lado, consideraron que estos niveles arenosos fueron depositados en un ambiente de dunas costeras. De todos modos, cualquiera de las dos interpretaciones indicaría muy leves desplazamientos de la línea de costa, ya sea por ligeros movimientos eustáticos, o por progradación de los sistemas costeros.
Luego, se depositaron uno o dos niveles de arenas con abundante micro y megafauna marina, equivalentes a los aflorantes en las costas de Diamante y posiblemente en todo el SO de Entre Ríos. En la parte superior se habría alcanzado el momento de mayor inundación, según se deduce de la composición y diversidad de la microfauna.
La sedimentación marina, o de transición, concluye con la depositación de un potente banco de pelitas verdes, ocasionalmente laminadas, con cristales de yeso muy bien desarrollados y concreciones calcáreas, y de un banco de arenas muy cementadas, con estructura granodecreciente.
Este conjunto señala la definitiva regresión del mar.
Por último se depositó un nuevo nivel de pelitas rojas masivas, con concreciones calcáreas, de origen indudablemente continental.
En resumen, la Formación. Paraná en la zona de Diamante, se depositó como resultado de una ingresión marina muy somera, regulada por la progradación de los ambientes costeros.
Hacia la parte superior de la Formación Paraná, el mar alcanzó su máximo ascenso y expansión, tras lo cual se retiró y se restableció la sedimentación netamente continental. Si consideramos a la Fm. Paraná como el conjunto de los depósitos marinos y de ambientes de transición, de tonalidades verdosas y amarillentas, claramente diferenciados de los depósitos rojizos de origen continental, posee una potencia de unos 60 metros, de los cuales sólo afloran unos 20-25 metros.
iii) La TEP al norte de la cuenca del Salado. Los depósitos de la TEP desde el norte de la cuenca del Salado, hasta sus manifestaciones más septentrionales conocidas, en el sur de Bolivia, poseen características bastante uniformes, sobre la base de sus caracteres litológicos y micropaleontológicos. Están formados, en todos los casos, por pelitas verdes masivas y areniscas verdes a amarillentas con un mayor o un menor contenido de fósiles marinos, y frecuentes niveles ricos en yeso.
En la mayoría de los perfiles se reconocen intercalaciones, de origen continental, de areniscas fluviales amarillas a grises, o de pelitas rojizas con yeso y concreciones calcáreas. Además, en casi todas las localidades, los depósitos marinos suprayacen y subyacen a depósitos de pelitas rojizas.
Los microfósiles marinos suelen encontrarse solamente en las areniscas verdes o amarillas con megafauna, siendo raros y muy mal preservados en las pelitas verdes. Las microfaunas son propias de ambientes marinos muy someros, generalmente hiposalinos. Los foraminíferos corresponden a la zona informal o hemerozona de P. tuberculatum (Malumián, 1970), con un marcado empobrecimiento en la diversidad y un notable incremento en la proporción de A. parkinsoniana, hacia el interior continental.
La temperatura del mar fue algo superior a la actual en los mares adyacentes, según los estudios de ostrácodos (Rossi de García, 1966; Zabert y Herbst, 1977; Zabert, 1978). Las faunas de foraminíferos son muy similares a las actuales de la plataforma argentina, excepto principalmente, por P. tuberculatum, especie extinta, y por D. bulbosa, actualmente restringida al norte del Brasil. La distribución actual de esta última especie apoyaría las conclusiones paleoclimáticas obtenidas de los ostrácodos.
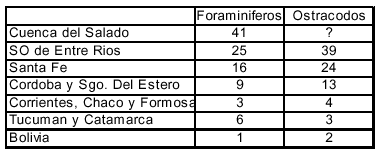
El mar ingresó desde el norte de la provincia de Buenos Aires, para luego extenderse hacia el centro y norte del país, y llegar hasta por lo menos, el sur de Bolivia y Paraguay (Figura 1), como indican el empobrecimiento de las microfaunas hacia el interior del continente (Cuadro 3) y la desaparición de las especies de salinidad normal, a partir de la provincia de Santa Fe. El límite oriental de la transgresión se ubicó al oeste del Uruguay y al SO de la provincia de Entre Ríos; el límite occidental está menos delimitado.
Ihering (1927), sobre la base de la notoria similitud entre las malacofaunas del mar Caribe y de la TEP, postuló la migración de algunos géneros de moluscos de aguas tropicales, desde el Caribe hacia la región del Plata, a través de un mar que habría cruzado el interior de América del Sur, de norte a sur, por la parte occidental del continente, y al que llamó la «Manga del Tethys» (Figura 3). Numerosos autores que estudiaron la TEP desde diferentes disciplinas, apoyaron la hipótesis de Ihering (Boltovskoy, 1958 y 1979, Boltovskoy y Lena, 1971 y 1974, Closs, 1962, del Río, 1990, y Pérez y Ramos, 1996, entre otros).
La distribución de los microfósiles de la TEP indica claramente que la migración de las faunas no se pudo producir a través de un mar intracontinental, (Cuadros 1 y 3), ya que el número de géneros disminuye rápidamente hacia el interior continental, conociéndose en Bolivia solamente un género de foraminífero, y dos géneros de ostrácodos de agua dulce. Por otro lado, hasta la fecha, no se hallaron las típicas faunas de moluscos afines a las del Caribe y del sur de Entre Ríos, más allá de la provincia de Santa Fe.
Es mucho más probable que las migraciones se hayan efectuado a través de la plataforma continental oriental de América del Sur, según fuera sugerido por Malumián (1970) y Sprechmann (1978).
Conclusiones
Se coincide con todos los trabajos micropaleontológicos previos, sobre las características paleoambientales y paleoecológicas de la TEP en la cuenca Chaco-Paranense y noroeste Argentino.
En términos generales fue un mar somero, hiposalino, y con una temperatura igual a algo superior a la actual de los mares adyacentes. No se hallaron elementos bioestratigráficos que permitan una datación más precisa de la TEP, por lo que se considera que se produjo en el período Mioceno medio-tardío.
La Formación Paraná, en la zona de Diamante, fue depositada por un mar somero, con intercalaciones de sedimentos de un ambiente de transición, probablemente deltaico, y no como producto de varios ciclos transgresivo-regresivos. La composición taxonómica de los microfósiles del subsuelo de la provincia de Entre Ríos, es muy similar a la conocida de afloramientos, excepto por algunos géneros y especies nuevas para la región. Sobre la base de la variación vertical de las microfaunas, se identificaron dos o tres períodos de inundación. El espesor de la Formación Paraná en la zona de Diamante es de unos 60 metros, considerando las facies marinas y transicionales.
Los foraminíferos depositados por la TEP en la cuenca Chaco-Paranense y el noroeste Ar-gentino, pertenecen a la zona informal o hemerozona de P. tuberculatum, con un marcado empobrecimiento de la diversidad hacia el interior continental, producto de la disminución de la profundidad y salinidad del mar. Sólo en cortos períodos se alcanzaron salinidades cercanas a las normales, en las provincias de Santa Fe y Entre Ríos. Sobre la base de la distribución geográfica de los microfósiles y moluscos se descarta su migración, desde el mar Caribe, a través del hipotético brazo marino intracontinental, conocido como la «Manga del Tethys»; la migración se habría efectuado a través de la plataforma continental oriental de América del Sur.
Agradecimientos
A las autoridades del SEGEMAR, por la cesión de los testigos de perforación; al Dr. Norberto. Malumián, por sus permanentes consejos; a la Lic. Adelma Bayarsky, por su desinteresada ayuda en el procesamiento del material; y a la Dra. Andrea Caramés, por la lectura crítica del manuscrito.
Apéndice: Sistemática
La sinonimia se restringe a la cita original, no se incluyen las citas de géneros con especie indeterminada. Los géneros de los foraminíferos según Loeblich y Tappan (1988). Para los géneros de los ostrácodos se mantienen los dados en los trabajos previos sobre la Formación Paraná.
(*)En estos trabajos se brindan descripciones e ilustraciones de material hallado en el país, preferentemente en la Formación Paraná.
Foraminíferos:
Textularia candeiana = Textularia candeiana d’Orbigny, 1839. En: de la Sagra. Hist. Phys. Polit. Natur. Cuba, p. 143, lám. 1, figs. 25-27. * Boltovskoy et al., 1980. Atlas, p. 51, lám. 32, figs. 8-11.
Textularia gramen = Textularia gramen d’Orbigny, 1846. Foram. Foss. Tert. Vienne, p. 248, lám. 15, figs. 4, 6. * Zabert y Herbst, 1977, FACENA, I, p. 146, lám. 1, fig. 3.
Cycloforina angulata = Miliolina bicornis var. angulata Williamson, 1858. Foram. Brit., p. 88, lám. 7, fig. 196. * Zabert y Barbano, 1984, FACENA, V, p. 143, lám. 1, fig. 1.
Pyrgo elongata = Biloculina elongata d’Orbigny, 1826. Ann. Sci. Nat. France, 1 (7), p. 298, n° 4. * Zabert, 1978, FACENA, II, p. 109, lám. 1, fig. 6.
Pyrgo patagonica = Biloculina patagonica d’Orbigny, 1839. Voy. Amér. Mérid., 5 (5), p. 65, lám. 3, figs. 15-17. * Zabert, 1978, FACENA, II, p. 108, lám. 1, fig. 5.
Pyrgo peruviana? = Biloculina peruviana d’Orbigny, 1839. Voy. Amér. Mérid., 5 (5), p. 68, lám. 9, figs. 1-3. * Boltovskoy et al., 1980. Atlas, p. 44, lám. 26, figs. 1-3.
Pyrgo ringens = Miliolites (ringens) subglobosa Lamarck, 1804. Ann. Mus. Nat. Hist., 5 (9), p. 351, lám. 17, fig. 1. * Boltovskoy et al., 1980. Atlas, p. 44, lám. 26, figs. 7-9.
Quinqueloculina sp. aff. Q. implexa = aff. Quinqueloculina implexa Terquen, 1886. Soc. Zool. France, Bull., 11, p. 335, lám 11, figs. 24-26. * Zabert, 1978, FACENA, II, p. 107, lám. 1, fig. 4.
Quinqueloculina bicornis = Serpula bicornis Walker y Jacob, 1798. Essays Microsc., p. 633, lám. 14, fig. 2. * Zabert, 1978, FACENA, II, p. 106, lám. 1, fig. 2.
Quinqueloculina lamarckiana = Quinqueloculina lammarckiana d´Orbigny, 1839. En: de la Sagra. Hist. Phys. Polit. Natur. Cuba, p. 164, lám. 11, figs. 14-15. * Zabert, 1978, FACENA, II, p. 107, lám. 1, fig. 3.
Quinqueloculina seminulina = Serpula seminulum Linnaeus, 1758. Systema Naturae, 1 (10), p. 786. * Zabert, 1978, FACENA, II, p. 106, lám. 1, fig. 1.
Ammonia compacta = Streblus compactus Hofker, 1964. p. 99, figs. 242-243. * Bertels y Zabert, 1980, 2° Congr. Arg. Paleo. y Bioest., Actas, 3, p. 71, lám. 1, figs. 5-6.
Ammonia parkinsoniana = Rosalina parkinsoniana d’Orbigny, 1839. En: de la Sagra. Hist. Phys. Polit. Natur. Cuba, p. 105, lám. 4, figs. 25-27. * Zabert, 1978, FACENA, II, p. 116, lám. 3, figs. 18a, b.
Bolivina costata = Bolivina costata d’Orbigny, 1839. Voy. Amér. Mérid., 5 (5), p. 62, lám. 8, figs. 8-9. * Zabert, 1978, FACENA, II, p. 112, lám. 2, fig. 12.
Bolivina sp. aff. B. robusta = aff. Bolivina robusta Brady, 1881. Quart. Jour., Micr. Sci., London, n. s., 21, p. 57. * Zabert, 1978, FACENA, II, p. 112, lám. 2, fig. 11.
Bolivina sp. cf. B. spathulata = cf. Textularia variabilis var. spathulata Williamson, 1858. Foram. Brit., p. 76, lám. 6, figs. 164-165. * Zabert, 1978, FACENA, II, p. 113, lám. 2, fig. 13.
Bolivina striatula = Bolivina striatula Cushman, 1922. Carnegie Inst. Washington, 311 (17), p. 27, lám. 3, fig. 10. * Zabert, 1978, FACENA, II, p. 111, lám. 2, fig. 10.
Buccella peruviana campsi = Eponides peruvianus campsi Boltovskoy, 1954. Inst. Nac. Inv. Cienc. Nat., Rev., Geol., 3 (3), p. 265, lám. 17, figs. 6-8.
B. peruviana frigida = Pulvinulina frigida Cushman, 1921 (1922). Canad. Biol. Fish., Contr., p. 12. * Zabert, 1978, FACENA, II, p. 115, lám. 2, fig. 16.
Bulimina affinis = Bulimina affinis d’Orbigny, 1839. En: de la Sagra. Hist. Phys. Polit. Natur. Cuba, p. 109, lám. 2, figs. 25-26. * Zabert, 1978, FACENA, II, p. 113, lám. 1, fig. 14.
Bulimina pseudoaffinis = Bulimina pseudoaffinia Kleinpell, 1938. Amer. Assoc. Petr. Geol., p. 257, lám. 9, fig. 9. * Zabert, 1978, FACENA, II, p. 114, lám. 2, fig. 15.
Buliminella elegantissima = Bulimina elegantissima d’Orbigny, 1839. Voy. Amér. Mérid., 5 (5), p. 51, lám. 7, figs. 13-14. * Zabert, 1978, FACENA, II, p. 110, lám. 1, fig. 9.
Cancris sagra = Rotalia sagra d’Orbigny, 1839. En: de la Sagra. Hist. Phys. Polit. Natur. Cuba, p. 91, lám. 5, figs. 14-15. * Zabert y Barbano, 1984, FACENA, V, p. 144, lám. 1, fig. 3.
Cibicides variabilis = Truncatulina variabilis d’Orbigny, 1839. Hist. Nat. Is. Canarias, p. 135, lám. 2, fig. 29. * Zabert y Herbst, 1977, FACENA, I, p. 145, lám. 1, fig. 9.
Cribroelphidium discoidale = Polystomella discoidale d’Orbigny, 1839. En: de la Sagra. Hist. Phys. Polit. Natur. Cuba, p. 76, lám. 6, figs. 23-24. * Zabert, 1978, FACENA, II, p. 117, lám. 3, fig. 19.
Disconorbis bulbosa = «Discorbis» bulbosa Parker, 1954. Mus. Comp. Zool. Harvard, Bull., 3 (10), p. 532, lám. 8, figs. 10-12. * Malumián, 1972, Ameghiniana, 9 (2), p. 106, lám. 6, fig. 2.
Fissurina bicarinata = Fissurina bicarinata Terquen, 1882. Soc. Géol. France, Mém., sér. 3, 2 (3), p. 31, figs. 24a, b. * Zabert, 1978, FACENA, II, p. 110, lám. 1, fig. 8.
Fissurina quadricostulata = Lagena quadricostulata Reuss, 1870. K. Akad. Wiss. Wien, Math.-Nat. Cl., 62 (1), p. 469, lám. 6, figs. 25-30. * Boltovskoy et al., 1980. Atlas, p. 32, lám. 16, figs. 5-7.
Globigerina juvenilis = Globigerina juvenilis Bolli, 1957. U. S. Nat. Mus., Bull., 215, p. 110, lám. 24, figs. 5a, b. * Zabert, 1978, FACENA, II, p. 118, lám. 3, fig. 21.
Globulina caribea = Globulina caribea d’Orbigny, 1839. En: de la Sagra. Hist. Phys. Polit. Natur. Cuba, p. 130, lám. 2, figs. 7-8. *f Zabert y Barbano, 1984, FACENA, V, p. 144, lám. 1, fig. 2.
Guttulina lactea = Serpula lactea Walker y Jacob, 1798. Essays Microsc., p. 634, lám. 14, fig. 4. * Zabert y Herbst, 1977, FACENA, I, p. 145, lám. 1, fig. 7.
Guttulina problema = Guttulina problema d’Orbigny, 1826. Ann. Sci. Nat. France, sér. 1, 7, p. 266, n° 14, fig. 61. * Malumián, 1972, Ameghiniana, 9 (2), p. 104, lám. 2, fig. 12.
Hanzawaia boueana = Rosalina bertheloti d’Orbigny, 1839. Hist. Nat. Is. Canarias, p. 135, lám. 1, figs. 28-30. * Zabert y Barbano, 1984, FACENA, V, p. 147, lám. 1, figs. 6a, b.
Nonion demens f. santamariana = Nonion demens f. santamariana Zabert, 1982. 3° Congr. Arg. Paleo. y Bioest., Actas, p. 185, lám. 1, figs. 1a-3b; lám 2, figs. 4a-7.
Nonionella atlantica = Nonionella atlantica Cushman, 1947. Cushman Lab. Foram. Res., Contr., 28, pt. 4, p. 90, lám. 20, figs. 4- 5. * Zabert y Barbano, 1984, FACENA, V, p. 146, lám. 1, fig. 5.
Nonionella auricula = Nonionella auricula Heron-Allen y Earland, 1930. Foram. Plymouth, p. 192, lám. 5, figs. 68-70. * Zabert y Barbano, 1984, FACENA, V, p. 145, lám. 1, fig. 4.
Nonionella auris = Valvulina auris d’Orbigny, 1839. Voy. Amér. Mérid., 5 (5), p. 47, lám. 2, figs. 15-17. * Zabert, 1978, FACENA, II, p. 119, lám. 3, fig. 22.
Protelphidium tuberculatum = Nonionina tuberculata d’Orbigny, 1846. Foram. Foss. Tert. Vienne, p. 108, lám. 5, figs. 13-14. * Zabert, 1978, FACENA, II, p. 117, lám. 3, fig. 20.
Rosalina floridana = Discorbis floridanus Cushman, 1922. Carnegie Inst. Washington, 311 (17), p. 39, lám. 5, figs. 11-12. * Zabert, 1978, FACENA, II, p. 115, lám. 2, fig. 17.
Ostrácodos:
Bensonia miocenica = Bensonia miocenica Rossi de García, 1969. Proc. 3rd Afr. Micropal. Coll. * Zabert, 1978. FACENA, II, p. 121, lám. 4, figs. 24a, b.
Bensonia reticulata = Bensonia reticulata Zabert, 1978. FACENA, II, p. 121, lám. 4, fig. 25; lám. 8, figs. 53a, b. Bradleya aff. prodictyonites = aff. Bradleya prodictyonites Benson, 1972. Smith. Contr. Paleobiol., 12. * Zabert y Herbst, 1977, FACENA, I, p. 154, lám. 2, fig. 3.
Buntonia entrerriensis = Buntonia entrerriensis Rossi de García, 1966. RAGA, 21 (3), p. 199, lám. 1, figs. 2a, b.
Callistocythere litoralensis = Perissocytheridea litoralensis Rossi de García, 1966. RAGA, 21 (3), p. 206, lám. 2, figs. 3a, b, c. Callistocythere marginalis = Callistocythere marginalis Zabert, 1978. FACENA, II, p. 134, lám. 6, fig. 40; lám. 9, figs. 61a, b, c, d.
Caudites kennedyi = Caudites kennedyi Rossi de García, 1966. RAGA, 21 (3), p. 203, lám. 3, figs. 2a, b.
Cornucoquimba lutziana = Cornucoquimba lutziana Zabert, 1978. FACENA, II, p. 143, lám. 7, fig. 48; lám. 8, figs. 58a, b, c, d.
Cyamocytheridea ovalis = Cyamocytheridea ovalis Rossi de García, 1966. RAGA, 21 (3), p. 203, lám. 4, figs. 4a, b.
Cyprideis camachoi = Cyprideis camachoi Rossi de García, 1966. RAGA, 21 (3), p. 202, lám. 1, fig. 5a.
Cyprideis herbsti = Cyprideis herbsti Bertels y Zabert, 1980. 2° Congr. Arg. Paleo. y Bioest., Actas, 3, p. 70, lám. 1, figs. 1a, b, 2a, b, 3.
Cytherella (C.) damotteae = Cytherella (Cytherelloidea) damotteae Rossi de García, 1966. RAGA, 21 (3), p. 207, lám. 1, figs. 1a, b.
Cytherella (C.) cf. C. (C.) punctata = cf. Cytherella (C.) punctata Brady, 1866. * Zabert, 1978, FACENA, II, p. 145, lám. 7, fig. 52.
Cytheretta argentinensis = Cytheretta argentinensis Rossi de García, 1966. RAGA, 21 (3), p. 206, lám. 2, figs. 4a, b.
Cytheretta punctata = Cytheretta punctata Sanguinetti, 1976. * Zabert, 1978, FACENA, II, p. 122, lám. 4, fig. 26.
Cytheropteron aff. newportense = aff. Cytheropteron newportense Crouch, 1949. * Rossi de García, 1966. RAGA, 21 (3), p. 205, lám. 4, figs. 2a, b, c.
Cytheropteron benedictus = Cytheropteron benedictus Rossi de García, 1966. RAGA, 21 (3), p. 204, lám. 4, figs. 1a, b, c.
Cytheropteron victoriensis = Cytheropteron victoriensis Rossi de García, 1966. RAGA, 21 (3), p. 204, lám. 2, figs. 2a, b, c.
Cytherura elegans = Cytherura elegans Zabert, 1978. FACENA, II, p. 129, lám. 5, figs. 32a, b; lám. 9, figs. 59a, b, c, d.
Cytherura rossiana = Cytherura rossiana Zabert, 1978. FACENA, II, p. 130, lám. 5, fig. 33; lám. 8, fig. 56.
Echinicythereis boltovskoyi = Echinicythereis boltovskoyi Rossi de García, 1966. RAGA, 21 (3), p. 202, lám. 2, figs. 1a, b.
Hemicytherura costulosa = Hemicytherura costulosa Zabert, 1978. FACENA, II, p. 131, lám. 6, figs. 36a, b; lám. 9, figs. 60a, b, c, d.
Henryhowella alwinae = Henryhowella alwinae Zabert, 1978. FACENA, II, p. 139, lám. 7, figs. 46a, b; lám. 10, figs. 67a, b, c, d.
Henryhowella. aff. evax = aff. Henryhowella evax (Ulrich y Bassler) Puri, 1957. * Rossi de García, 1966. RAGA, 21 (3), p. 200, lám. 1, figs. 3a, b, c.
Loxoconcha paranensis = Loxoconcha paranensis Rossi de García, 1966. RAGA, 21 (3), p. 203, lám. 1, fig. 4a.
Munseyella argentina = Munseyella argentina Zabert, 1978. FACENA, II, p. 137, lám. 7, figs. 43a, b; lám. 10, figs. 65a, b, c.
Murrayina grekoffi = Murrayina grekoffi Rossi de García, 1966. RAGA, 21 (3), p. 201, lám. 3, figs. 3a, b, c, d, e.
Paijenborchella punctacostata = Paijenborchella punctacostata Zabert, 1978. FACENA, II, p. 138, lám. 7, fig. 44; lám. 10, figs. 66a, b, c, d.
Paijenborchella aff. punctacostata = aff. Paijenborchella punctacostata Zabert, 1978. FACENA, II, p. 138, lám. 7, fig. 44; lám. 10, figs. 66a, b, c, d.
Paracytheridea? laudata = Paracytheridea? laudata Rossi de García, 1966. RAGA, 21 (3), p. 205, lám. 3, figs. 1a, b.
Patagonacythere anzoteguiana = Patagonacythere anzoteguiana Zabert, 1978. FACENA, II, p. 142, lám. 7, fig. 49; lám. 10, figs. 68a, b, c.
Patagonacythere paranensis = Patagonacythere paranensis Zabert, 1978. FACENA, II, p. 140, lám. 7, fig. 47; lám. 9, figs. 63a, b, c, d, e.
Pellucistoma santafesinensis = Pellucistoma santafesinensis Zabert, 1978. FACENA, II, p. 136, lám. 6, figs. 42a, b; lám. 9, figs. 62a, b, c, d.
Perissocytheridea alvareziana = Perissocytheridea alvareziana Zabert, 1978. FACENA, II, p. 126, lám. 5, fig. 30; lám. 8, fig. 56.
Perissocytheridea ornata = Perissocytheridea ornata Zabert, 1978. FACENA, II, p. 127, lám. 5, figs. 35a, b; lám. 8, figs. 55a, b, c.
Perissocytheridea victoriensis = Perissocytheridea victoriensis Zabert, 1978. FACENA, II, p. 125, lám. 5, figs. 29a, b; lám. 8, figs. 54a, b, c.
Platella fragilis = ?Platella fragilis Rossi de García, 1966. RAGA, 21 (3), p. 207, lám. 3, fig. 4a.
Pumilocytheridea herbsti = Pumilocytheridea herbsti Zabert, 1978. FACENA, II, p. 123, lám. 5, figs. 34a, b; lám. 10, figs. 64a, b, c.
Trachyleberis nova = Trachyleberis nova Rossi de García, 1966. RAGA, 21 (3), p. 200, lám. 4, figs. 3a, b.
Wichmanella deliae = Wichmanella deliae Bertels, 1975. Ameghiniana, 12 (3), p. 272, lám. 4, figs. 3-4. * Zabert y Herbst, 1977, FACENA, I, p. 159, lám. 3, fig. 5.
Bibliografía
Aceñolaza, F. G. y Aceñolaza, G. F. 1999. Trazas fósiles del Terciario marino de Entre Ríos (Formación Paraná, Mioceno medio), República Argentina. Academia Nacional de Ciencias (Córdoba), Anales 64: 209-233.
Becker, D. y Bertels, A., 1980. Micropaleontología de la secuencia terciárica de la perforación Puelche (margen continental argentino). II Congreso Argentino de Paleontología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología (Buenos Aires, 1978), Actas 2: 315-333.
Bertels, A. y Zabert, L. L., 1980. Microfauna del Grupo Santa María (Terciario Superior) en las Provincias de Catamarca y Tucumán, República Argentina. II Congreso Argentino de Paleontología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología (Buenos Aires, 1978), Actas 3: 67-73.
Boltovskoy, E., 1958. The foraminiferal fauna of the Río de la Plata and its relation to the Caribbean area. Contributions from the Cushman Foundation for Foraminiferal Research, 9: 17-21.
Boltovskoy, E., 1979. Paleoceanografía del Atlántico sudoccidental desde el Mioceno, según estudios foraminiferológicos. Ameghiniana, 16 (3-4): 357-389.
Boltovskoy E., 1980. Perforación Gil 1, provincia de Buenos Aires (foraminíferos, edad, paleoambiente). Ameghiniana, 17 (4) 339-362.
Boltovskoy, E. y Lena, H., 1971. The Foraminifera (except family Allogromiidae) wich dwell in fresh water. Journal of Foraminiferal Research, 1 (2): 71-76.
Boltovskoy, E. y Lena, H., 1974. Foraminíferos del Río de la Plata, Argentina. Servicio de Hidrología Naval, H 661: 1-22.
Boltovskoy, E., Giussani, G., Watanabe, S., and Wright, R., 1980. Atlas of Benthic Shelf Foraminifera of the Southwest Atlantic. Dr. W. bv Publishers, The Hague-Boston-London: 58 p., 36 lám., 17 figs.
Closs, D., 1962. Foraminiferos e Tecamebas de Lagoa dos Patos (R. G. S.). Escola de Geología da Universidade Federal do Rio Grande do Sul (Porto Alegre), Boletín. 11: 1-130.
Del Río, C. J., 1990. Comparación, origen y significado paleoclimático de la Malacofauna entrerriense. Academia Nacional de Ciencias Exactas, Físicas y Naturales (Buenos Aires), Anales 42: 205-223.
Frenguelli, J., 1920. Contribución al conocimiento de la Geología de Entre Ríos. Academia Nacional de Ciencias (Córdoba), Boletín 24, Entregas 1 y 2.
Gavriloff, I. J. C., Durango de Cabrera, J. y Vergel, M. M., 1998. Paleontología de invertebrados, paleobotánica y palinología. En: Gianfrancisco, M., Puchulu, M. E., Durango de Cabrera, J. y Aceñolaza, G. F. (Eds.), Geología de Tucumán, Publicación Especial del Colegio de Graduados en Ciencias Geológicas de Tucumán, pp. 211-226.
Herbst, R. y Zabert, L. L., 1987. Microfaunas de la Formación Paraná (Mioceno superior) de la cuenca Chaco-Paranense (Argentina). Facultad de Ciencias Exactas y Naturales del Litoral, Revista 7: 165-206.
Ihering, H., 1927. Die Geschiechte des Atlantischen Ozeans. Gustav Fischer Verlag, Jena: 237 p. Loeblich, A. R., Jr. y Tappan, H., 1988. Foraminiferal genera and their classification. Van Nostrand Reinhold, New York, New York: 970 p., 847 lám. Malumián, N., 1970. Bioestratigrafía del Terciario marino del subsuelo de la Provincia de Buenos Aires (Argentina). Ameghiniana, 7 (2): 173-204.
Malumián, N., 1972. Foraminíferos del Oligoceno y Mioceno del subsuelo de la Provincia de Buenos Aires. Ameghiniana, 9 (2): 97-137.
Malumián, N., 1978. Esbozo paleoecológico de las asociaciones foraminiferológicas terciarias de la Argentina. Ameghiniana, 15 (1-2): 161-171.
Malumián, N., Suriano, J. M. y Cobos, J. C., 1998. La Formación Barranca Final en su localidad tipo. Mioceno, cuenca del Colorado. X Congreso Latinoamericano de Geología y VI Congreso Nacional de Geología Económica (Buenos Aires, 1998), Actas 1: 125-130. Marshall, L. G., Sempere, T. y Gayet, M., 1993. The Petaca (late Oligocene-middle Miocene) and Yecua (late Miocene) Formations of the Subandean-Chaco Basin, Bolivia, and their tectonic significance. Docum. Lab. Géol. Lyon, 125: 291-301, 2 figs., 1 lám.
Masiuk, V., Becker, D. y García Espiasse, A., 1976. Micropaleontología y sedimentología del pozo YPF.Ch.PV.es-1 (Península Valdés) Provincia del Chubut, República Argentina. Importancia y Correlaciones. Arpel XXIV, Yacimientos Petrolíferos Fiscales (Buenos Aires), 22 pp.
Murray, J. W., 1991. Ecology and Palaeoecology of Benthic Foraminifera. Longman Scientific & Technical, Essex, England: 397 p. Pérez, D. J. y Ramos, V. A., 1996. Los depósitos sinorogénicos. En: V. A. Ramos et al. (Eds.), Geología de la Región del Aconcagua, provincias de San Juan y Mendoza, Subsecretaría de Minería de la Nación, Dirección Nacional de Servicio Geológico, Anales, 24 (11): 317-341.
Pisetta, J. L., 1968. [Descripción de una faunula de Foraminíferos de la Provincia de Entre Ríos. Trabajo Final de Licenciatura, Universidad de Buenos Aires, Inédito].
Rossi de García, E., 1966. Contribución al conocimiento de los ostrácodos de la Argentina. I, Formación Entre Ríos, de Victoria, Provincia de Entre Ríos. Asociación Geológica Argentina, Revista 21, 3: 94-208.
Rossi de García, E., 1969. Algunos ostrácodos del Entrerriense de Paraná, Provincia de Entre Ríos, República Argentina. Asociación Geológica Argentina, Revista 24, 3: 276-280.
Sprechmann, P., 1978. The Paleoecology and Paleogeography of the Uruguayan Coastal Area During the Neogene and Quaternary. Reprinted from Zitteliana Abhandlungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie (München) 4: 3-72, 6 lám.
Stappenbeck. R., 1926. [Geologie und Grundwasserkunde der Pampa. Dirección Nacional de Minería y Geología, Traducción del original, 1943 (Buenos Aires), inédito].
Uliana, M. A. y Biddle, K. T., 1988. Mesozoic-Cenozoic Paleogeographic and Geodynamic evolution of Southern South America. Revista Brasileira de Geociências 18 (2): 172-190.
Windhausen, A., 1931. Geología Argentina. Segunda parte: Geología histórica y regional del territorio argentino. Editorial Peuser, Buenos Aires, 646 pp.
Zabert, L. L., 1978. Micropaleontología de la Formación Paraná (Mioceno Superior) en el subsuelo de la Provincia de Santa Fe, República Argentina. Facultad de Ciencias Exactas y Naturales del Litoral, Revista II: 101-165.
Zabert, L. L., 1984. Nonion demens (Bik), foraminífero bentónico en el Mioceno del Valle de Santa María (Tucumán) y Selva (Santiago del Estero), Argentina. III Congreso Argentino de Paleontología y Bioestratigrafía (Corrientes, 1982), Actas: 183-196.
Zabert, L. L. y Barbano, J., 1984. Microfósiles miocénicos (Formación Paraná) de Córdoba, Santiago del Estero y nuevas descripciones de Entre Ríos. Facultad de Ciencias Exactas y Naturales del Litoral, Revista 5: 135-150.
Zabert, L. L. y Herbst, R., 1977. Revisión de la microfauna Miocena de la Formación Paraná (entre Victoria y Villa Urquiza, Provincia de Entre Ríos) con algunas consideraciones estratigráficas. Facultad de Ciencias Exactas y Naturales del Litoral, Revista 1: 131-174.
Recibido: 20 de abril de 2000
Aceptado: 1 de noviembre de 2000