Análisis de la vegetación en tres tipos de hábitat de Carpinchos en la baja cuenca del Río Paraná, Argentina
Rubén D. QUINTANA1; Nora MADANES1; Ana I. MALVÁREZ1; Fabio A. KALESNIK1y Marcela CAGNONI1
1Departamento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Buenos Aires, 1428 Capital Federal; Argentina. E-mail: rubenq@bg.fcen.uba.ar
Abstract: COVER_TYPES CHARACTERIZATION IN THREE CAPIBARA’S HABITAT IN THE LOWER BASIN OF THE PARANA RIVER, ARGENTINA. The objective of the present work was identify and characterize the different cover-types in three capybara's habitat types of the lower basin of the Paraná River (Bajo Delta, Puerto Constanza and Villaguay) throughout the vegetation features. The different cover-types were defined according to the covering and dominance of species as well as the relative topographic position. Specific composition and abundance of the plant species were estimated in each of the different cover-types. These data were analyzed using a Correspondence Analysis (AC) in each habitat types. The whole floristic composition of the three areas was compared using a Sperman's Rank Correlation Coefficient. Results showed that the vegetation of Bajo Delta and Puerto Constanza was related with variations of the topographic position and, therefore, the flooding levels. In the Villaguay, on the other hand, vegetation in each cover-type was the results of the tree density and grazing level. Floristic composition was a adequate indicator of the natural and antropic differences within and among the different habitat types. Relative topographic position, flooding regime and disturbance level would be the main factors affecting the vegetation structure in the different covertypes of the three-studied habitat types.
Key words: Correspondence Analysis - Cover-types - Capybara - Habitat - Paraná River - Argentina.
Palabras clave: Análisis de Correspondencia - Ambientes - Capibara - Hábitat - Río Paraná - Argentina.
Introducción
Para gran parte de la fauna silvestre, la vegetación es la variable que otorga las características estructurales del hábitat en los ambientes terrestres (Rotenberry y Wiens 1980). En particular, para muchas especies, la fisonomía de la vegetación ha sido reconocida como un factor de gran importancia a escala de paisaje en la determinación de su distribución. Por una parte esta influye en la selección de hábitat y por otra en la disponibilidad de recursos críticos tales como alimento, sitios de reproducción, nidificación o refugio de depredadores (Hildén 1965; Wiens 1992). En particular, el carpincho es un herbívoro que dado sus hábitos anfibios pastorea en la cercanía de los cuerpos de agua (Escobar y González Jiménez 1976; Jorgenson 1986, Quintana 1996). Tiene cuatro requerimientos esenciales: disponibilidad de agua, forraje y áreas para descanso y refugio, los que determinarán la proporción de hábitat disponible (Ojasti 1973; Herrera y Macdonald 1989). Además, la configuración espacial de los ambientes que conforman su hábitat será un factor de importancia en su distribución, puesto que utiliza las interfases tierra-agua, descartando las áreas alejadas a estas interfases y las zonas internas de los grandes cuerpos de agua (Quintana 1999).
Clements (1928) planteó que las especies vegetales podían ser consideradas como un indicador de la presencia de determinadas especies animales, dado que el tipo de vegetación representaría el producto de las principales condiciones ambientales presentes (van der Maarel 1979). Es así que, tanto por la información que aporta la vegetación per se, como por las inferencias que pueden hacerse sobre otros factores ambientales, la misma resulta una variable de importancia en cualquier estudio sobre fauna silvestre.
Por otra parte, la herbivoría ejerce su efecto sobre la vegetación de forma tal que su estructura es el resultado de la interacción de las poblaciones vegetales y las especies herbívoras, ya sea naturales o introducidas (Sala 1988; Quintana 1996).
Cuando el objetivo de un estudio es analizar el patrón de distribución de una especie es recomendable comenzar por identificar y analizar cuantitativamente las principales características del hábitat (Ben-Shahar y Skinner 1988). Por lo tanto, el objetivo del presente trabajo ha sido identificar y caracterizar los diferentes ambientes presentes en tres tipos de hábitat de carpinchos (Hydrochaeris hydrochaeris) en la baja cuenca del Río Paraná a partir de los atributos estructurales de la vegetación.
Esta caracterización a nivel de ambientes constituyó la base para análisis posteriores sobre selección y uso del hábitat por parte de los carpinchos (Quintana 1996).
Dada su extensión, esta cuenca posee una diversidad de regiones con distintas características biogeográficas. Se parte de la premisa de que el clima, la topografía y el patrón de inundabilidad, entre otras variables, condicionan la distribución de comunidades y especies definiendo la aptitud de hábitat para este roedor.
Materiales y Métodos
AREA DE ESTUDIO
La Cuenca del Plata es considerada por su extensión, tamaño y caudal, la segunda en importancia
en Sudamérica. Comprende en su parte inferior las llanuras costeras del sur de Entre Ríos y del
noroeste bonaerense, sectores costeros aislados de la República Oriental del Uruguay y el delta del
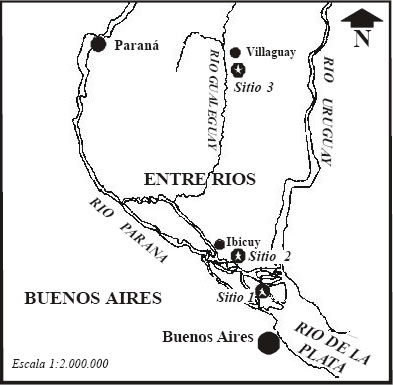
Paraná. Para el presente trabajo se seleccionaron tres áreas: Bajo Delta, Puerto Constanza y Villaguay.
Las dos primeras se encuentran ubicadas en la región del Delta del Río Paraná (33° 36' y 34° 26' S y 58°
00' y 59° 30' O; (Fig. 1). Esta constituye una compleja planicie inundable definida como un extenso
macromosaico de humedales (Malvárez 1999). El clima es templado subhúmedo con lluvias todo el
año. Los valores medios de temperatura, humedad relativa y precipitación para el período 1981-1990
fueron de 17,0 º C, 76% y 1087, 1 mm, respectivamente (estación meteorológica "San Pedro INTA";
Servicio Meteorológico Nacional, 1992).Desde el punto de vista biogeográfico, es considerada como
una ingresión subtropical en el distrito pampeano. Esto determina la coexistencia de especies propias
de ambas zonas, dando al área un perfil característico (Ringuelet 1961)
En la porción entrerriana las principales actividades productivas son la ganadería extensiva, la apicultura y la recolección de leña en tanto que en la porción bonaerense se desarrolla la forestación con salicáceas y el turismo. Estas actividades, conjuntamente con las obras de infraestructura para el manejo del agua, han provocado, al menos en algunos sectores, cambios significativos en la abundancia y distribución espacial de muchas especies vegetales y animales (Kandus 1997; Kalesnik y Malvárez 1996; Quintana et al. 2002).
El Bajo Delta es la porción más austral de esta región (33° 48' y 34o 26' S y 59° 00' y 58° 31' O) presenta una morfología típicamente deltaica (Malvárez 1999). Por sus características ecológicas constituye un humedal sometido a mareas de agua dulce cuyo régimen hidrológico está determinado por las inundaciones periódicas de los ríos Paraná y de la Plata y en menor medida de los ríos Uruguay y Gualeguay (Kandus 1997). Las islas presentan un perfil de tipo cubeta con una zona perimetral alta o "albardón" y, una porción interior deprimida (80% de su superficie). En el albardón se asentaban bosques ribereños (Burkart 1957) que fueron reemplazados principalmente por plantaciones de álamos (Populus sp.). En la porción interior predominan bosques abiertos de ceibos y pajonales que representan las únicas comunidades naturales actuales (Kandus y Adámoli 1993) aunque parte de éstos han sido drenados y forestados con sauces (Salix sp.). En la actualidad, la forestación es la actividad productiva más importante tanto desde el punto de vista económico como por su extensión e impacto ecológico (Kandus 1997). En consecuencia, el paisaje está constituido por un mosaico de forestaciones activas y abandonadas, terraplenes, zanjas y canales agropecuarios conjuntamente con parches de los pajonales y ceibales.
El área de estudio se ubica en una forestación localizada en la intersección de los ríos Paraná Guazú y Paraná Miní, con una superficie de 8000 ha (Fig. 1)
Fig. 1. Ubicación de los tres sitios estudiados en la Baja Cuenca del Río Paraná
Puerto Constanza (33°48'30"S, 58°58'76"O; Fig. 1) se ubica en un área transicional entre el Bajo Delta y la zona de "bosques, praderas y arroyos de cordones y depresiones" (Malvárez 1999). El paisaje se caracteriza por la presencia alternada de cordones arenosos y zonas de medias lomas y de bajos. Las primeras presentan anegamiento temporario y los bajos anegamiento temporario ó permanente. Paralelos a los cordones se encuentran cauces lénticos, en general cegados e incluso interrumpidos por la actividad antrópica. El régimen hidrológico está principalmente determinado por el río Paraná Guazú, que aporta aguas en los picos de crecida en forma de flujos mantiformes (Malvárez 1999). Este, conjuntamente con el escaso microrrelieve local determinan una predominancia de bañados. Los ambientes de alto presentan una fisonomía de bosque abierto o arbustal, cuya especie dominante es el espinillo (Acacia caven). Las media lomas constituyen praderas con predominio de herbáceas graminiformes y equisetoides, y latifoliadas típicas de suelos sujetos a inundación. Cuando la media loma es suficientemente amplia se diferencia una porción más elevada de composición similar a los estratos bajos del bosque y una más baja con especies herbáceas graminiformes típicas de ambientes que se inundan con frecuencia. En las partes más deprimidas de los bajos predominan juncales (Schaenoplectus californicus). En algunos casos, las zonas centrales de estas áreas deprimidas presentan agua libre. Los cauces, en general, se encuentran cubiertos por especies acuáticas flotantes y arraigadas (Quintana 1996).
La principal actividad productiva es la ganadería extensiva, que en el caso del campo estudiado (de aproximadamente 700 ha.) su carga animal fue de 0,7 vacas/ha, existiendo también algunas zonas forestadas con salicáceas.
Villaguay se ubica en las tierras altas de Entre Ríos, 25 km al sur de la ciudad homónima (31° 34' 30" S, 59° 01' 02" O; Fig. 1). El paisaje presenta un relieve suavemente ondulado cuyo río principal es el Gualeguay. El clima es templado, y de rasgos subtropicales, con temperaturas y precipitaciones abundantes en primavera y verano decreciendo de noreste a sudoeste. Los valores medios de temperatura, humedad relativa y precipitación para el período 1981-1990 fueron de 18,2 º C, 76% y 1110,5 mm, respectivamente (Servicio Meteorológico Nacional 1992). Fitogeográficamente, pertenece al distrito del ñandubay de la Provincia del Espinal (Cabrera 1971). Las actividades más desarrolladas son la agricultura y la ganadería mixta, con predominio de invernada. El campo estudiado (de aproximadamente 600 ha) está dedicado principalmente a la ganadería, con algunas zonas de cultivo. Está atravesado por el arroyo Segovia, caracterizado por la presencia de agua libre en la mayor parte de su curso, con pequeños parches de Polygonum punctatumr, Pistia stratiotes, Azolla filiculoides y ciperáceas, bordeado por un bosque en galería empobrecido e invadido por especies exóticas como el paraíso (Melia azedarach) y el ligustro (Ligustrum lucidum). Estos relictos se encuentran cercados impidiendo el ingreso del ganado. El resto del área presenta una fisonomía de sabana abierta y se encuentra intensamente pastoreada tanto por ganado bovino y ovino como por carpinchos.
A diferencia de Puerto Constanza la carga animal fue muy alta, encontrándose en el orden de las 3 vacas/ha y 6 ovejas/ha.
ANÁLISIS DE LA VEGETACIÓN
El estudio se llevó a cabo durante las primaveras de 1991 y 1992 en los tres sitios descriptos
anteriormente. Cada sitio fue estratificado mediante el análisis de fotografías aéreas en escala 1:20.000,
con posterior corroboración e interpretación a campo de los ambientes naturales y antrópicos identificados.
Estos fueron definidos de forma tal que dieran como resultado una división general del área basada en la fisonomía y en la posición topográfica relativa (altos, medias lomas y bajos). En la
tabla 1 se presentan los ambientes identificados para cada sitio.
En cada sitio se realizaron censos ubicados al azar sobre transectas orientadas según el principal gradiente topográfico local. En Bajo Delta se realizaron muestreos en cuadrados de 2x2 m, donde se tomaron datos de abundancia-cobertura de especies según la escala de Braun-Blanquet modificada (Mueller-Dombois y Ellemberg 1974). Para Puerto Constanza y Villaguay se empleó el método del punto de intercepción (Hays et al. 1981), empleando transectas de 10 m con intervalos de muestreo cada 20 cm, lo que equivalió a un total de 50 puntos por transecta.
En el Bajo Delta se realizaron 262 censos, en Puerto Constanza 46 y en Villaguay 59. El tamaño de la muestra por ambiente estuvo en relación con la heterogeneidad interna de los mismos y con su superficie, estimándolo a través del grado de fluctuación de la media (Matteucci y Colma 1982).
Las especies vegetales fueron determinadas a campo con posterior corroboración en laboratorio. Para su identificación se siguió a Cabrera y Zardini (1993) y Burkart (1974).
ANÁLISIS DE DATOS
La vegetación de los 3 sitios fue comparada a través del coeficiente de correlación por rangos de
Spearman (Zar 1996). Para el análisis de la vegetación de cada sitio se empleó un análisis de correspondencia,
AC (Gauch et al. 1977; Kent y Coker 1992). Los datos de cobertura y abundanciacobertura
de las especies fueron transformados según la escala propuesta por van der Maarel (1979).
La diversidad de especies vegetales para cada sitio y en cada ambiente se calculó mediante el índice de
Shannon-Weaver (H') (Magurran 1991). Complementariamente se analizaron la riqueza florística y la dominancia, como el logaritmo del número de especies presentes en la muestra (R) y como el índice
de dominancia (H = H'/R), respectivamente (Magurran 1991).
Los ambientes estrictamente acuáticos, si bien fueron caracterizados en función de su vegetación, no fueron considerados para el presente análisis.
Resultados
ANÁLISIS A ESCALA REGIONAL
El índice de correlación de Spearman mostró diferencias en la vegetación a escala regional. La
comparación entre localidades dio como resultado una correlación negativa, siendo mayor para las
comparaciones entre Villaguay y los sitios deltaicos (rs =-0,39, N= 234; p<0,00001 y rs =-0,44, N=203; p<0,00001, para la comparación de Villaguay con Bajo Delta y Puerto Constanza, respectivamente).
La comparación entre Bajo Delta y Puerto Constanza mostró una mayor afinidad entre las
localidades deltaicas (rs =-0,17, N= 194; p=0,021).
ANÁLISIS A ESCALA LOCAL
Bajo Delta. Se identificaron 8 ambientes terrestres además de las zanjas y canales agropecuarios
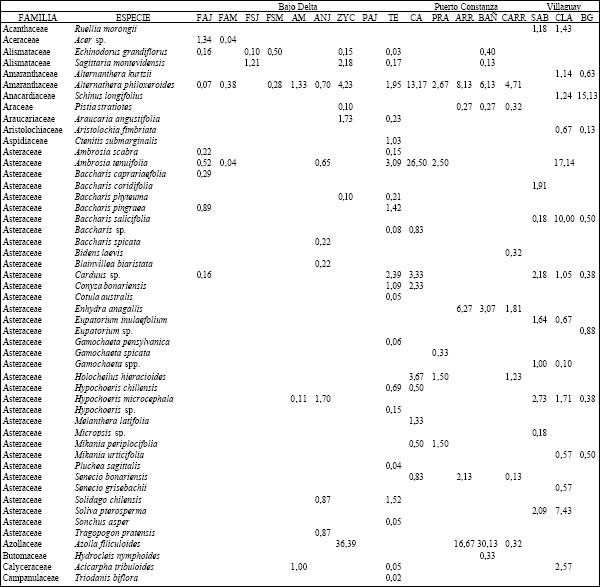
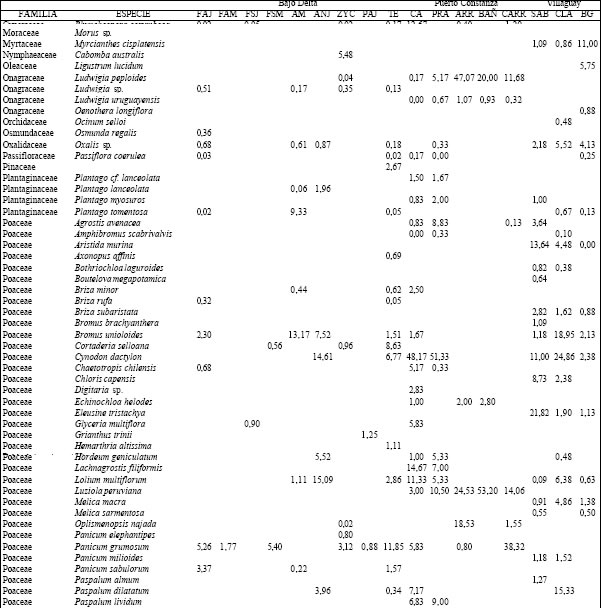
(Tabla 1), registrándose un total 157 especies cuyos valores de cobertura-abundancia porcentuales (o
valor de importancia) se presentan en la Tabla 2. Las comunidades vegetales de estos ambientes
difirieron en su fisonomía y composición específica. Así, en las forestaciones, las principales especies
del sotobosque y del estrato arbustivo fueron Carex riparia, Rubus sp. y Scirpus giganteus aunque con
distinto valor de importancia (Tabla 2). En los ambientes de alto C. riparia presentó valores mayores
al 24 %, en tanto que en los de bajo fueron del orden del 12 %. La situación inversa la presentó S.
giganteus que en las forestaciones de sauce joven (FSJ) se registró con un valor superior al 45 %
mientras que en los altos o estuvo ausente o solo se registró con valores cercanos al 3 %. La estructura
del sotobosque para los bosques de acacia negra joven (ANJ) y de acacia negra madura (ANM) fueron
diferentes entre sí y respecto a los de otros tipos de bosques. Mientras que en el pajonal (PAJ)
dominó S. giganteus, en los terraplenes (TERR) Panicum grumosum fue la especie dominante en el
sotobosque mientras que Populus sp. Constituyó la especie más abundante en el estrato alto. Las
zanjas y los canales agropecuarios se caracterizaron por la presencia de Azolla filiculoides.
En la Tabla 1 se observa que el PAJ y la FSJ presentaron la menor diversidad y una baja equitatividad. En las forestaciones de sauce maduro (FSM) y en las forestaciones de álamo maduro (FAM), por el contrario, los bajos valores de diversidad se relacionan tanto con bajos valores de equitatividad y de riqueza. La situación opuesta la presentaron los TERR y las forestaciones de álamo joven (FAJ) con una alta diversidad producto de la alta equitatividad (baja dominacia) y de una muy alta riqueza .
En la figura 2 se observa la distribución de censos resultado del AC. La varianza explicada por el primer y cuarto eje fue del 53 % (autovalor l= 0,83 y 0,52 respectivamente) Hacia el extremo positivo del primer eje se segregan los censos dominados por S. giganteus pertenecientes a los ambientes de bajo de los PAJ y de las FSJ, y hacia el negativo los censos pertenecientes a los ambientes de alto de los ANM y los ANJ, dominados por G. triacanthos. Sobre el cuarto eje, se segregan en el extremo positivo los censos dominados por G. triacanthos y Salix sp. pertenecientes a los ambientes ANM y FSM. Estos ambientes presentan un estrato arbóreo denso (>60%) mientras que en el sotobosque no se observa ninguna especie dominante (con valores de importancia mayores al 20%). La composición del sotobosque es la variable que diferencia a estos dos ambientes (Tabla 2). En el extremo negativo se segregan los censos con un estrato arbóreo abierto correspondientes al TERR (líneas de árboles plantadas a lo largo de algunos terraplenes) y un estrato herbáceo codominado por S. giganteus, C. riparia, Cortaderia selloana y Panicum grumosum las cuales se disponen a lo largo del microgradiente observado en estos ambientes.
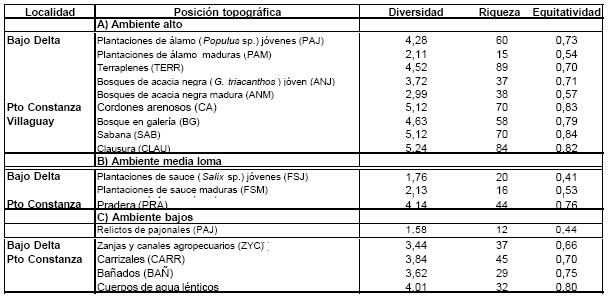
Tabla 1. Ambientes presentes en las localidades estudiadas,de acuerdo a su posición topográfica : A) ambientes de alto, B) de media loma y C) de bajo. Para cada ambiente se indican los valores de diversidad, riqueza y equitatividad
 |
 |
 |
 |
 |
 |
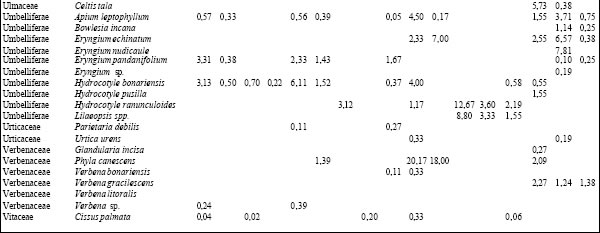 |
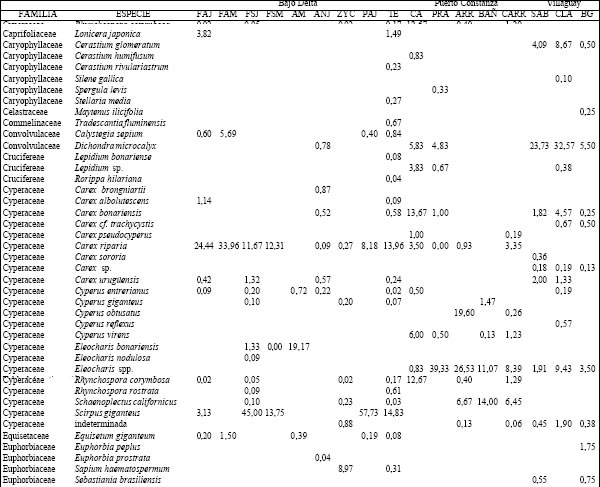
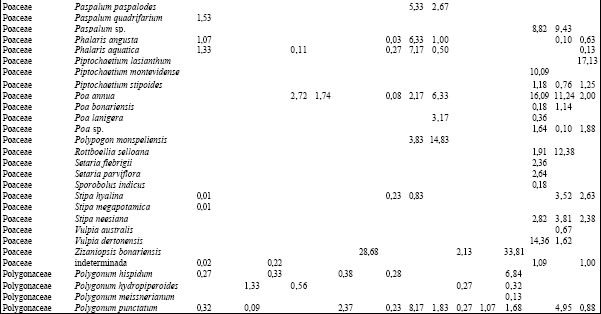
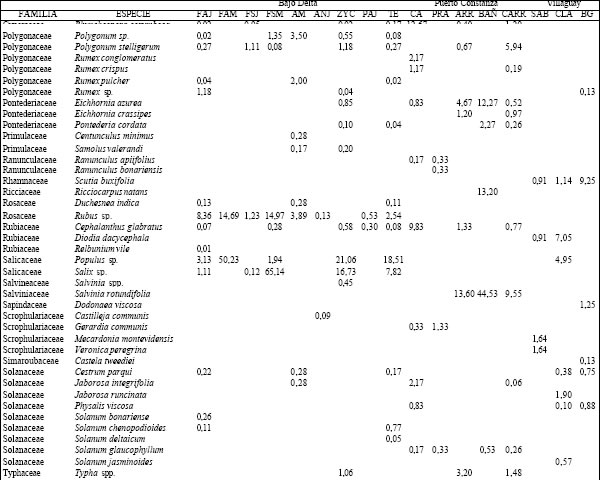
Tabla 2. Composición florística porcentual de las diferentes especies vegetales en cada uno de los ambientes presentes en los distintos sitios.TAER= Tierras altas de Entre Ríos. FAJ= forestación de "Alamo" joven; FAM= forestación de "Alamo" maduro; FSJ = forestación de "Sauce" joven; FSM = forestación de "Sauce" maduro; AM = bosque de "Acacia negra" madura; ANJ = bosque de "Acacia negra" joven; ZYC = zanjas y canales agropecuarios; PAJ= pajonal; TE= terraplenes; CA= cordón arenoso; PRA= pradera; ARR= arroyo; BAÑ= bañado; CARR: carrizal; SAB= sabana; CLA= clausura; BG= bosque en galería.
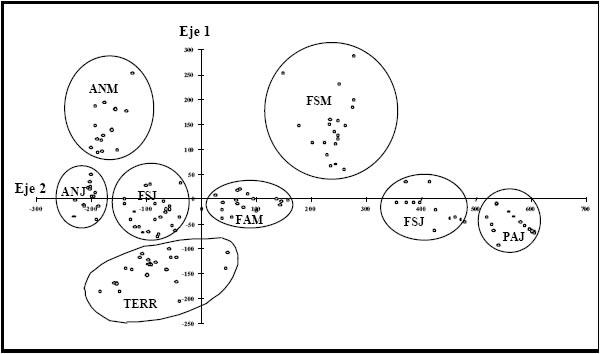
Fig. 2. Análisis de correspondencia (CA) basado en datos de abundancia de especies vegetales tal como se observa en el anexo 1 para Bajo Delta. Gráfico bidimensional entre los valores del primer y cuarto eje. FAM= forestación de álamo maduro; FAJ= forestación álamo joven; ANM= bosque maduro de acacia negra; ANJ= bosque joven de acacia negra; TERR= terraplenes; FSM= forestación de sauce maduro; FSJ= forestación de sauce joven; PAJ= pajonal.
Puerto Constanza. Se identificaron 5 ambientes caracterizados en función de la permanencia del agua (Tabla 1): los cordones arenosos (CA) mostraron una fisonomía pluriestratificada en tanto que el resto de los ambientes presentaron un solo estrato; éstos se diferenciaron también en su composición florística. Se registraron 154 especies cuyos valores de abundancia-cobertura se presentan en la Tabla 2. Así, en el estrato arbóreo de los CA dominó A. caven aunque con bajo valor de importancia (10%) y en el herbáceo C. dactylon al igual que en la pradera (PRA). En el arroyo (ARR) dominó Ludwigia peploides acompañada por Limobium laegivatum mientras que L. peruviana y Salvinia rotundifolia presentaron los mayores valores de importancia en el bañado (BAÑ) y P. grumosum en el CARR.
En la Tabla 1 se observa que todas las comunidades presentaron una alta diversidad aunque dentro de este sitio el BAÑ y el CARR fueron los ambientes menos diversos. Un rasgo importante a destacar son los altos valores de equitatividad observados para todos los ambientes. La riqueza fue mínima en el BAÑ y en el ARR, explicando la menor diversidad, y máxima en el CA, lo que, junto con la mayor equitatividad, explicaría el alto valor de diversidad observado en este ambiente.
En la figura 3a se observa la distribución de censos resultado del AC. La varianza explicada por los dos primeros ejes fue del 63% (autovalor l=0,86 y 0,75, respectivamente). Hacia el extremo positivo del primer eje se segregan los censos caracterizados por la presencia de P. grumosum (38,2%) que definen el carrizal y lo separan del resto de los ambientes. Esta separación resulta clara puesto que el carrizal constituye una comunidad altamente homogénea. Sobre el eje 2 se separan los ambientes de acuerdo al gradiente topográfico local: en el extremo positivo se agrupan los censos correspondientes a los ambientes de bajo (BAÑ, dominados por L. peruviana (53,2%) y CARR. El BAÑ quedó definido por un mosaico de parches caracterizados por 2 grupos de censos caracterizados por S. californicus por una parte y el resto de las acuáticas flotantes y arraigadas por la otra. En la parte positiva del eje 2 se observa también un grupo de censos que definen el ecotono pradera-bañado, caracterizados por la presencia de especies comunes a PRA y BAÑ, particularmente Eleocharis spp. y gramíneas bajas como L. peruviana. Hacia el negativo se segregan los censos pertenecientes a los ambientes CA y PRA dominados por C. dactylon con un valor de importancia mayor al 48%. Cuando se graficó conjuntamente el primer eje con el cuarto eje del AC (Fig. 3b), se observa una separación de los censos de la PRA de aquellos del AC (autovalor l= 0,33; 13% de la varianza total explicada).
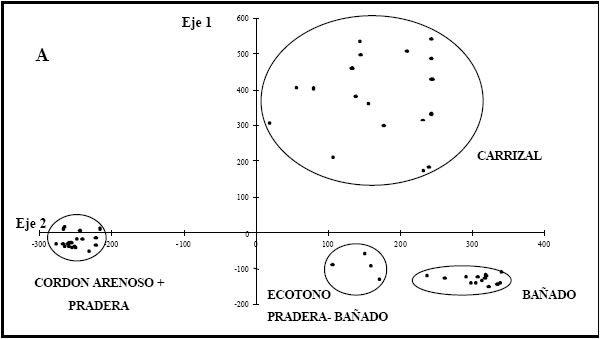
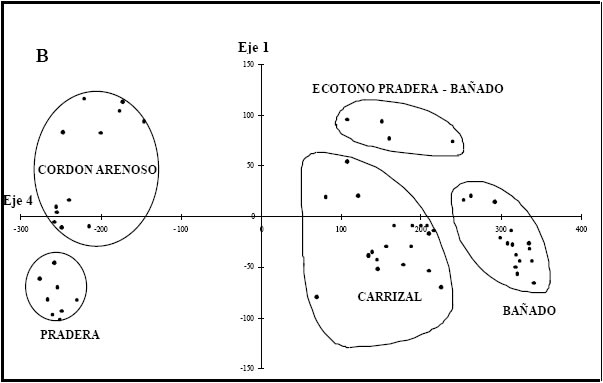
Fig. 3. Análisis de correspondencia (CA) basado en datos de abundancia de especies vegetales tal como se observa en la Tabla 1 para Pto. Constanza. a) Gráfico bidimensional entre los valores obtenidos para el primer y segundo eje. b) idem a) pero entre los valores obtenidos para el primer y cuarto eje.
Villaguay. Se identificaron tres tipos de ambientes: bosque en galería (BG), sabana abierta pastoreada (SAP) y, dentro de ésta, una clausura (CLA) vedada al ingreso del ganado doméstico durante más de 30 años (Tabla 1). A diferencia de los sitios anteriores, los tres ambientes se diferenciaron por el tipo de disturbio al que estuvieron o están sometidos, registrándose un total de 122 especies cuyos valores de abundancia cobertura porcentuales se presentan en la Tabla 2. Las comunidades vegetales difirieron en su fisonomía y composición cuali y cuantitativa. Así en la SAP y en la CLA se observó una codominancia de Dichondra microcalix (23,7% y 32,6%, para SAP y CLA, respectivamente) con Eleusine tristachya (21,8%) en el primer caso y con C. dactylon (24,9%) en el segundo. En el estrato arbóreo de estos ambientes se observó la presencia de A. caven con un valor de importancia inferior al 12%. El BG se caracterizó por la presencia de un estrato arbóreo codominado por Schinus longuifolia, Melia azedarach y Myrcianthes cisplatensis (con valores entre 11 y 15%) y por alta dominancia de Piptochaetium lastianthum (17%) en el sotobosque.
En la Tabla 1 se observa que todas las comunidades presentaron una alta diversidad (aunque esta fue menor para el BG) y una alta equitatividad. La riqueza fue mínima en el BG y máxima en la CLA, lo cual explicaría las diferencias observada en los valores de diversidad.
En la Fig. 4 se observa la distribución de censos resultado del AC. La varianza explicada por los dos primeros ejes fue del 69 % (autovalor l=0,67 y 0,36 respectivamente). Sobre el primer eje, se segregan en el extremo positivo los censos de la SAP y en el negativo los de la CLA. Si bien la especie dominante es la misma (D. microcalyx), la composición específica de aquellas especies que presentan valores intermedios de importancia es diferente, lo cual estaría reflejándose en la separación de dichos grupos. La disposición de estos dos grupos a lo largo de este eje estaría indicando un gradiente de disturbio asociado al efecto de la herbivoría.
Hacia el extremo positivo del segundo eje, por otra parte, se segregan los censos pertenecientes al BG y hacia el extremo negativo los censos pertenecientes a los ambientes SAP y CLA, caracterizados por la presencia de un bosque abierto. Sobre este eje, los ambientes se dispusieron en función de un gradiente de intensidad lumínica dado por la estructura de la vegetación leñosa.
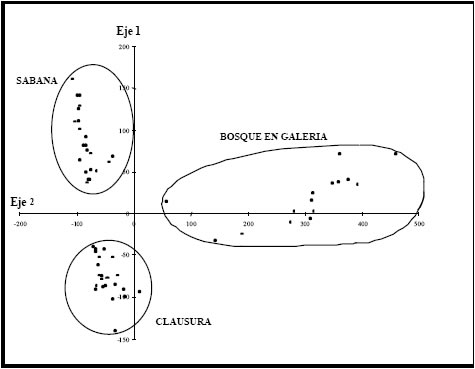
Fig. 4. Análisis de correspondencia (CA) basado en datos de abundancia de especies vegetales tal como se observa en la Tabla 1 para Villaguay. Gráfico bidimensional entre los valores obtenidos para el primer y segundo eje.
Discusión
ANÁLISIS A ESCALA REGIONAL
En el presente estudio, tal como fuera señalado por otros autores (Matteucci y Colma 1982; ter
Braak 1993), la estructura de la vegetación resultó ser un buen indicador de las diferencias en las
variables tanto naturales como antrópicas que caracterizaron las distintas áreas y los distintos ambientes
que conforman los paisajes de cada una de ellas. Bajo Delta y Puerto Constanza mostraron
una mayor afinidad en la vegetación dada por la presencia de un gradiente de inundabilidad que
condiciona la existencia de zonas de bajo con una gran abundancia de especies hidrófilas, gran parte
de ellas compartidas. Dicha similitud se mantiene a pesar de la transformación que ha sufrido el Bajo
Delta por la actividad forestal. En Villaguay, por el contrario, no se observa un perfil topográfico
marcado y el paisaje se encuentra además muy afectado por la ganadería. Tanto las diferencias observadas
en el régimen hidrológico, la topografía y la actividad humana entre las localidades deltaicas y
Villaguay dan como resultado una expresión característica diferencial de la vegetación.
ANÁLISIS A ESCALA LOCAL - LOCALIDADES DELTAICAS.
En ambas localidades, la comunidad vegetal que caracterizó a cada uno de los ambientes identificados
respondió en una primera instancia a la permanencia del agua o inundabilidad, relacionada
con la posición en el gradiente topográfico local. Esto se observó aún en el caso del Bajo Delta, a
pesar de la profunda transformación del paisaje original por actividades humanas. En este sentido,
la capacidad de las plantas de sobrevivir y germinar en los humedales de agua dulce está condicionada
por las características particulares del régimen hidrológico. Esto determina diferencias en la composición
específica de los distintos ambientes (Whigham y Simpson 1977; Leck y Simpson 1987). El
hidroperíodo y la energía del agua son factores reguladores primordiales en estos humedales y
funcionan como factores favorables o desfavorables para las plantas (Van der Valk y Welling 1988;
Lugo et al. 1990; Brinson 1993 a; 1993b). Su acción da como resultado la conformación de un tamiz
ambiental para la instalación y persistencia de las especies (Van der Valk 1981). En ambas localidades
es factible encontrar tipos de vegetación adaptados a zonas de alto, media loma y bajo, determinados
por la interacción de la posición topográfica y del régimen hidrológico.
En el Bajo Delta, además, se manifiesta también un gradiente lumínico condicionado por la estructura del dosel de los distintos tipos de bosques presentes.
Bajo Delta. Con respecto a los ambientes presentes en este sitio, los terraplenes ocuparon las posiciones más elevadas dentro del microrrelieve local y pudiéndose encontrar sin forestar o con baja densidad de árboles, presentando buen drenaje y gran intensidad lumínica. El bosque de ANM, ubicado en el antiguo albardón de un arroyo y en la media loma, se caracterizó por una alta densidad de árboles y por un canopeo cerrado, siendo el sotobosque ralo, con especies adaptadas a condiciones de baja luminosidad El bosque ANJ es la consecuencia de la invasión de esta especie sobre una antigua pista de aterrizaje abandonada construida por relleno de un área inicialmente deprimida. Esta pista presentó una vegetación de pastizal de composición similar a la de los pastizales continentales de la Pcia. de Buenos Aires con renovales de acacia negra dispersos (Cabrera 1976).
La baja diversidad y riqueza de los ambientes de bajo, cuya situación extrema correspondió al pajonal de S. giganteus, es un atributo de las comunidades naturales del Bajo Delta, las que se caracterizan por ser simples y por estar dominadas por una pocas especies (Kandus et al. 2003).
A pesar de la transformación del Bajo Delta, los ambientes antrópicos presentan abundantes graminoides (sensu Barkman 1988) características de la vegetación natural de esta región, tales como C. riparia, S. giganteus y P. grumosum, entre otras. C. riparia, en particular, resultó más abundante en ambientes antropizados que en los parches de pajonal natural. Esta característica se relacionaría con su alta capacidad de adaptabilidad a aquellos ambientes que se encuentran sometidos a disturbios antrópicos (Kandus et al. 2000). P. grumosum, presente en TERR y distintas forestaciones, ha sido citada como invasora (Burkart 1957). Además de estas especies nativas, cabe mencionar la presencia de malezas típicas de los campos altos de Buenos Aires y Entre Ríos como C. dactylon y Lolium multiflorum en ANJ y esta primera y Bromus unioloides en TERR, las que son comunes en ambientes disturbados.
Dentro de las especies invasoras presentes, la acacia negra, además de encontrarse en el Bajo Delta se la observa en la ribera del Río de la Plata (Morello et al. 2000). El carácter invasor se debe a su rápido desarrollo (Ruiz Selmo 1998) y a la dispersión de semillas realizada por aves(de Urquiza 1999), que en el sitio de estudio, por cotorras comunes –(Myopsitta monacha). Dos especies invasoras arbustivas presentes en el sotobosque de ambientes forestados son la madreselva (Lonicera japonica), de origen asiático y presente solo en la FAJ y la zarzamora (Rubus sp.), de origen europeo y que también se dispersa por acción de las aves. Esta última llega a invadir desde la forestaciones las zonas periféricas del pajonal formando matorrales muy densos.
Puerto Constanza. A diferencia del Bajo Delta, en este sitio los ambientes identificados dentro del gradiente topográfico son de origen natural, con una baja modificación dada por el pastoreo extensivo.
Las comunidades vegetales presentaron una predominancia de gramíneas y latifoliadas, tal como fuera observado por Malvárez (1997) y Franceschi et al. (1979, 1985) en ambientes similares de la baja cuenca del río Paraná. Esta característica marca una diferencia importante con respecto al Bajo Delta, donde predominan las ciperáceas. Además, otras especies presentes en ambas localidades mostraron diferencias importantes en sus valores de abundancia. Así, mientras que C. riparia y S. giganteus fueron muy abundantes en Bajo Delta, en esta localidad ambas son raras.
En los CA la especie característica fue A. caven, la cual, cabe destacar que ésta es una especie xerofítica lo suficientemente plástica como para adaptarse a estos humedales y soportar períodos de anegamiento más o menos prolongados. Es de amplia distribución en el Chaco, el Monte, el Espinal y la Provincia Paranaense, donde se comporta como invasora (Demaio et al. 2002).
Los bajos, por otra parte, se caracterizaron por la presencia de amplios bañados con gramíneas hidrófilas tales como L. peruviana , Echinochloa helodes y Oplismenopsis najada, las cuales se encuentran ausentes o muy poco representadas en el Bajo Delta. Otra especie abundante en estos bañados es S. californicus, poco presente en la zona estudiada del Bajo Delta. Sin embargo, la porción distal de esta región presenta grandes extensiones de esta especie, tal como fuera observado por Kandus et al. (2003). P. grumosum aparece en esta localidad como dominante en ambientes de bajo lo cual resulta contrastante con lo observado en el Bajo Delta, donde se encuentra principalmente en ambiente antropizados de alto y bajo. Malvárez (1997) observó que los albardones de las islas de la zona de Victoria (Entre Ríos) están dominados por especies del género Panicum (P. prionites, P. grumosum y P. rivulare). Estas observaciones muestran que esta especie es lo suficientemente plástica como para adaptarse a ambientes con diferente grado de anegamiento.
Por último, si bien el CA y la PRA se corresponden con fisonomías diferentes, en ambos domina C. dactylon y además presentan una gran similitud cualitativa de herbáceas bajas comunes, aunque con distinto valor de importancia. Esto se debe a que, por una parte la media loma alta y el CA casi no difieren desde el punto de vista topográfico y a que el estrato arbóreo es lo suficientemente abierto como para no generar un gradiente lumínico.
Villaguay. Este sitio se diferenció de las localidades deltaicas por escasa presencia de especies hidrófilas y la mayor riqueza de leñosas nativas. Otra diferencia observada fue la ausencia de un gradiente topográfico marcado y de un régimen de inundaciones periódico. En este caso las diferencias observadas entre ambientes estuvieron dadas por la acción del pastoreo del ganado doméstico combinado con un gradiente lumínico producto de la densidad diferencial de leñosas. Esta diferencia se debe, por una parte, a la extracción selectiva de madera que tuvo lugar en el pasado y por otro, a la ausencia de renovales por efecto de la herbivoría en la actualidad. En este caso, el efecto de la herbivoría potenciaría al de la tala ya que la primera estaría impidiendo la regeneración de leñosas por consumo directo de los renovales. Esto explicaría la falta de bosque en galería en gran parte de la costa del arroyo Segovia y de leñosas en algunos sitios de la sabana. Así, la preservación de los relictos de bosque en galería como los de la clausura ha sido posible debido a su aislamiento del impacto del pastoreo por ganado.
Además, la alta carga animal presente en este sitio habría contribuido a alterar la estructura y composición florística de la vegetación, tal como ha sido señalado por otros autores (McNaughton 1983; Weller 1981; McNaughton y Sabuini 1988; Sala 1988).
Por otra parte, la invasión de leñosas exóticas (ligustro y paraíso) en los parches relictuales del bosque en galería promueven cambios en la estructura del mismo. Sin embargo, este bosque a pesar de estar empobrecido, presenta una alta diversidad y riqueza de especies. Su cerrada canopia impide el desarrollo de una abundante cobertura de especies del sotobosque, aunque el mismo está constituido por un importante número de especies herbáceas cuya dominante (P. lasianthum) se encuentra ausente en los otros ambientes.
En cuanto al efecto de la presión de pastoreo sobre la vegetación, Sala (1988) observó que el material verde se concentraba fundamentalmente entre los 0 y 5 cm de altura en áreas pastoreadas mientras que en las clausuras no pastoreadas éste se concentró entre los 10 y 30 cm. Si bien en este estudio no se cuantificó la biomasa verde, se observó que en la SAP, la cual estaba muy pastoreada, la altura de la vegetación no superaba los 5 cm mientras que en la clausura, éste fue superior a los 20 cm, dando como resultado una estructura de herbáceas diferente al área intensamente pastoreada.
Por otra parte se observaron diferencias en la composición y abundancia de especies y en la riqueza específica entre estos ambientes pero los valores de diversidad y equitatividad fueron similares. La riqueza no solo fue menor en la SAP, sino que en la CLA se observó un mayor número de especies latifoliadas. Así, el pastoreo influyó de manera diferencial entre ambos sitios favoreciendo o desfavoreciendo el desarrollo de ciertas especies, lo cual se puede observar al ver los valores de importancia de algunas de ellas como Bromus unioloides, Eleusine trystachia y Ambrosia tenuifolia, entre otras.
Sala (1988) señala que los pastizales húmedos y subhúmedos con una corta historia evolutiva de pastoreo son los más susceptibles a ser alterados por la herbivoría y que la diversidad de especies declina rápidamente a medida que la intensidad de pastoreo se incrementa. En nuestro caso, la menor intensidad de herbivoría en la CLA (solo pastoreada por carpinchos) se expresó en una alta riqueza de especies y una mayor densidad de leñosas y altura del estrato herbáceo. La CLA fue el único ambiente donde pudo constatarse la presencia de Senecio grisebachii Back y una mayor riqueza y abundancia de especies del género Eryngium L. Este hecho difiere de lo planteado por Cabrera (1971), quien señala que la primer especie se ha hecho muy abundante en la región debido a su baja palatabilidad para el ganado mientras que Eryngium aparecería asociado a áreas sometidas a sobrepastoreo. Por otra parte, B. coridifolia sólo estuvo presente en la SAP, la cual es evitada por el ganado por su toxicidad.
Conclusión
La metodología empleada en este estudio permitió clasificar, definir y caracterizar los ambientes en estas tres localidades, identificando los factores naturales y antrópicos determinantes del tipo de vegetación presente. Los resultados aquí obtenidos conjuntamente con la información disponible sobre los requerimientos de hábitat del carpincho constituyen la base para el planteo de modelos de la relación carpincho-hábitat para estas tres localidades.
Agradecimientos
Queremos expresar nuestra gratitud a los Sres. Juan Pita y Oscar Maglione y a la familia Jauregui por permitirnos realizar el presente estudio en sus respectivos establecimientos. A las familias González y Cívico y al Sr. Alfredo Pulenta por la ayuda brindada en el campo. Un agradecimiento especial a R. Vicari por la lectura crítica del trabajo. El presente trabajo fue realizado con los subsidios EX 135 y Ex 143 del Programa UBACyT de la Universidad de Buenos Aires.
Bibliografía
Ben-Sahar, R. y J. Skinner. 1988. Habitat preference of african ungulates derived by uni and multivariate análisis. Ecology 69: 1479-1485.
Braun-Blanquet, J. 1979. Fitosociología. H. Blume, Madrid, 547 pp.
Brinson, M. 1993a. A Hydrogeomorphic Classificationfor Wetlands. Wetland Research Program TechnicalReport WRPDE-4. US Army Corps of Engeneers,Waterways Experiment Station. Washington, D.C.
Brinson, M.. 1993b. Changes in the Functioning of Wetlands along Environmental Gradients. Wetlands, vol 13, No 2 pp 65-74.
Burkart, A. 1957. Ojeada sinóptica sobre la vegetación del Delta del Río Paraná. Darwiniana 11: 457-561.
Burkart, A 1969. Flora ilustrada de Entre Ríos (ARGENTINA). Parte II. Colección Científica INTA. Tomo VI, II. Buenos Aires, 551 pp.
Burkart, A 1974. Flora ilustrada de Entre Ríos (ARGENTINA). Parte VI. Colección Científica INTA, Buenos Aires, 554 pp.
Burkart, A 1979. Flora ilustrada de Entre Ríos (ARGENTINA). Parte V. Colección Científica INTA, Buenos Aires, 550 pp.
Burkart, A 1987. Flora ilustrada de Entre Ríos (ARGENTINA). Tomo VI, Parte III. Colección Científica INTA. Buenos Aires, 763 pp.
Cabrera, A. 1971. Fitogeografía de la República Argentina. Bol. Soc. Argent. Bot.14: 1-42.
Cabrera, A y E. M. Zardini. 1993. Manual de la flora de los alrededores de Buenos Aires. ACME, Buenos Aires, 755 pp.
Clements, F. 1928. Plant succession and indicators. H.W. Wilson Co., New York, 453 pp.
Demaio, P., Karlin, U.O. y Medina, M. 2002. Árboles nativos del centro de Argentina. L.O.L.A., Buenos Aires. 127 pp.
De Urquiza, M. 1999. Análisis comparativo del crecimiento de una especie leñosa nativa y otro exótico de los talares de Magdalena.Tesis de Licenciatura, Universidad de Buenos Aires, Buenos Aires, 78 pp.
Escobar, A. y González Jiménez, E. 1976. Estudio de la competencia alimenticia de los herbívoros mayores del llano inundable con especial referencia al chigüire (Hydrochaeris hydrochaeris). Agr. Tropical 26: 215-227.
Franceschi, E.A y Lewis J.P. 1979. Notas sobre la vegetación del valle santafesino del Río Paraná (República Argentina). Ecosur (6)11: 55-82
Franceschi, E.A, Prado D y Lewis J.P. 1985. Comunidades vegetales y mapa de vegetación de la Reserva “El Rico” e islas aledañas (Provincia de Santa Fe, Rep. Argentina). Servicio de publicaciones UNR (Universidad Nacional de Rosario). 41 pp
Gauch, H. G., R. H. Whittaker and R. T. Wentworth. 1977. A comparative study of non-metric ordinations. J. of Ecol.69: 135-152.
Hays, R. L., C. Summers and W. Seitz. 1981. Estimating wildlife habitat variables. Technical Report FWS/OBS-81-47, USDI, Fish and Wildlife Service, Washington D.C.
Herrera, E. and D.W. Macdonald. 1989. Resource utilization and territoriality in group-living capybaras (Hydrochoerus hydrochaeris). Journal of Animal Ecology 58: 667-679.
Hildén, O. 1965. Habitat selection in birds. Ann. Zool. Fenn. 2: 53-75.
Instituto Nacional de Tecnología Agropecuaria. 1973. El Delta del Paraná. Rev. EEA INTA Delta del Paraná 14. 110 pp.
Jorgenson, J. 1986. Notes on the ecology and behaviour of capybaras in Northeartern Columbia. Vida Silvestre Neotropical 1: 31-40.
Kandus, P. 1997. Análisis de los patrones de vegetación a escala regional en el Bajo Delta Bonaerense del Río Paraná (Argentina).Tesis doctoral, Universidad de Buenos Aires, Buenos Aires, 241 pp.
Kandus, P. y J. Adámoli. 1993. Freshwater marsh vegetation response to flooding dynamic in the Lower Delta of Paraná River. Wetlands Ecol. and Manage. 2(4): 213-222.
Kandus, P., F. Kalesnik, L. Borgo y A. I. Málvarez. 2001. La reserva Natural “Isla Botija” en el Delta del Río Paraná: análisis de las comunidades de plantas y condicionantes ambientales. Parodiana 12 (en prensa).
Kandus, P., Málvarez , A.I. y N Madanes.2003. Estudio de las comunidades de plantas herbáceas de las islas del Bajo Delta del río Paraná. Darwiniana 41(1-4): 00-000. 2003
Kent, M. and P. Coker. 1992. Vegetation description and analysis. A practical approach. CRC Press, Boca Raton, Ann Arbor, 363 pp.
Malvárez, A. I. 1997. Las comunidades vegetales del Delta del Río Paraná. Su relación con factores ambientales. Tesis Doctoral, Universidad de Buenos Aires, Buenos Aires, 167 pp.
Mc Naughton, S.J. and G.A. Sabrini.1988. Large african mammals as regulators of vegetation structure: 339-354. En: M. J. A. Werger, P. J. M. van der Aart, H. J. During &. J. T. A. Verhoeven (eds.) Plant form and vegetation structure. SPB Academic Publishing, The Hague.
Matteucci, S. y A. Colma. 1982. Metodología para el estudio de la vegetación. Secretaría Gral. de la OEA. Progr. Regional de Desarrollo Científico y Tecnológico. Monogr. Nº 22, Ser. Biología, Washington D.C., 168 pp.
Morello, J., G. D. Buzai, C. A. Baxendale, S.D. Matteucci, A. F. Rodríguez, R. E. Godagnone y R. R. Casas. 2000. Urbanización y consumo de Tierra fértil. Ciencia Hoy 10: 50-61.
Ojasti, J. 1973. Estudio biológico del chigüire o capibara. Fondo Nacional de Investigaciones Agropecuarias, Caracas, 275 pp.
Quintana, R. D. 1996. Análisis y evaluación de la aptitud de hábitat del carpincho (Hydrochaeris hydrochaeris) en relación con la heterogeneidad del paisaje y las interacciones con ganado doméstico. Tesis doctoral, Universidad de Buenos Aires, Buenos Aires, 270 pp.
Quintana, R.D. 1999. Relación entre la estructura del paisaje en un humedal y la fauna silvestre: el carpincho (Hydrochaeris hydrochaeris) como caso de estudio. En: Pp. 185-204 Tópicos sobre humedales subtropicales y templados de Sudamérica (Malvárez, A. I., ed.). ORCyT – MAB/ UNESCO, Montevideo.
Ringuelet, R. 1961. Rasgos fundamentales de la zoogeografía de la Argentina. Physis 22: 150-170.
Rotenberry , J. T. and J.A. Wiens. 1980. Habitat structure, patchiness, and avian community in North American steppe vegetation: A multivariate analysis. Ecology 61: 1228-1250.
Ruiz Selmo, F. 1998. Estudio ecofisiológico de Gleditsia triacanthos. Comparación entre una especie leñosa invasora y una nativa en los talares de Magdalena. Tesis de Licenciatura, Universidad de Buenos Aires, Buenos Aires, 80 pp.
Sala, O. E. 1988. The effect of herbivory on vegetation structure: 317-330. En M. J. A. Werger, P. J. M. van der Aart, H. J. During &. J. T. A. Verhoeven (eds.) Plant form and vegetation structure. SPB Academic Publishing, The Hague.
Servicio Meteorológico Nacional 1992. Estadísticas climatológicas 1981-1990, Ser. B6, Nº 37, 1ª ed., Fuerza Aérea Argentina, Buenos Aires, 704 pp.
Ter Braak, C. J. F. 1993. Ordination: 91-173. In R. H. G. Jongman, C. J. F. ter Braak & O. F. R. van Tongeren (eds). Data analysis in community and landscape ecology. Pudoc, Wageningen, 299 pp.
Van der Maarel, E. 1979. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 39: 97-114.
Wiens, J. A. 1992. Ecology of bird communities, Vol. 1-2. Cambridge Univ. Press, Cambridge.
Zar, J. 1984. Biostatistical analysis. 2nd Edition Prentice-Hall Inc., New Jersey, 718 pp.