Integridad ecológica en los ríos Paraná y Mississippi: ¿trayectorias paralelas o divergentes?
Claudio BAIGÚN1; Norberto OLDANI2 y John NESTLER3
1Instituto Tecnológico de Chascomús, (IIB-INTECH), CONICET-Universidad Nacional de General San Martín,
Camino de Circunvalación Laguna Km 6, C.C. 164 (7130) Chascomús, Pcia. de Buenos Aires, Argentina.
E-mail: claudiobaigun@intech.gov.ar
2Instituto de Desarrollo Tecnológico para la Industria Química. INTEC, Güemes 3450, 3000 Santa Fe, Argentina.
E-mail: gbio@ceride.gov.ar
3Environmental Laboratory, U.S. Army Engineer Research and Development Center Vicksburg, MS 39180-6199.
Trabajo presentado en el Seminario Internacional sobre Manejo de Humedales en América Latina y Simposio
Internacional de Ecoturismo y Humedales. Paraná (E.R), Argentina, 25 al 27 de set 2003.
Abstract: ECOLOGICAL INTEGRITY IN THE PARANA AND MISSISSIPI: ¿PARALLEL OR DIVERGENT TRAGETORY? Large floodplain rivers form aggregated ecosystems of singular value and unique complexity due to the integration of biochemical, geomorphological, hydrological and biological processes taking place at different spatial and temporal scales. This study compares the main characteristics of the Superior Mississippi and Middle Paraná as leading cases of opposed conservation and management scenarios. In the Superior Mississippi geomorphology has been altered by the construction of impoundments (pools); hydrology was modified by channel training structures and most of alluvial floodplains were lost due to levees construction. At present the river is severely fragmented by dams built to improve navigation conditions. Also changes of hydrological pulses have modified biogeochemical cycles affecting inputs of nutrients, particulated and dissolved organic matter and sediments to floodplains. On the other hand the Middle Paraná still exhibits a complex alluvial valley where biochemical cycles are strongly linked to carbon metabolism and floodplain community biodiversity is associated to annual hydrological pulses. This case represents a model of a lowly disturbed floodplain river providing important clues for rehabilitation, restoration and remediation projects in other similar systems. Examples of such opposed scenarios reflect the different conditions found in large floodplain rivers allowing to predict if trajectories will preserve or reduce the ecological integrity of such systems
Key words: Ecology - Río Paraná - Río Mississippi
Palabras clave: Ecologia - Río Paraná - Río Mississippi.
Introducción
Los grandes ríos con llanura de inundación, constituyen ecosistemas agregados de valor singular y complejidad única por la interrelación y variedad de procesos biogeoquímicos, geomorfológicos, hidrológicos, biológicos y ecológicos que se manifiestan a diferentes escalas espaciales y temporales. Gran parte de estos cursos de agua, tienen modificados significativamente sus características ecológicas y los patrones estructurales y funcionales, por alteración de los cauces principales y la desaparición de cauces secundarios, asociado al desarrollo de hidrovías, la utilización de su llanura aluvial para agricultura, pérdida de la vegetación riparia (por obras costeras), alteraciones de los hábitat por represamientos (principalmente para navegación, generación de electricidad y riego). Un caso notable y paradigmático como la falta de perspectiva del uso de los recursos, condujo a un estado de degradación severa la integridad ecológica, lo constituye el río Mississippi en América del Norte. Por el contrario, otros ríos como el Paraná en América del Sur aún retienen sus características naturales y representan excelentes ejemplos de complejidad.
El objetivo del trabajo es comparar las características más importantes de dos grandes ríos con planicies de inundación: el Mississippi Superior y el Paraná Medio como casos testigo y paradigmas de condiciones opuestas de conservación y manejo.
El Mississippi: Ejemplo de la degradación de un río único.
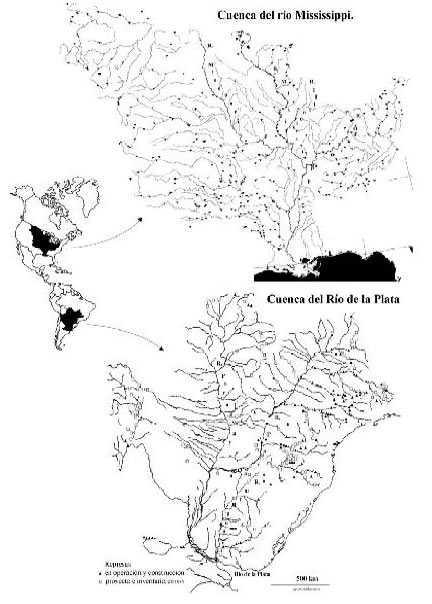
El río Mississippi constituye el mayor río de América del Norte. Con una cuenca de 3.267 Km2 y una longitud de 3.730 Km, atraviesa el continente en sentido norte-sur desde el lago Itaska en Minnesota hasta Louisiana, donde se anastomosa y desemboca en el golfo de México (Figura 1).
Fig. 1. Ubicación geográfica de las cuencas de los ríos Mississippi y Paraná
El Mississippi sufrió importantes modificaciones y alteraciones. En sus riberas se levantaron 3.500 Km de terraplenes que ocupan 12.300 Km2, 330 Km de espigones y 1.370 Km de empalizadas. Esto, directa o indirectamente favoreció la pérdida de humedales costeros, llanuras aluviales y la reducción de la intensidad de los pulsos de inundación. La rectificación del cauce, para favorecer la navegación, eliminó meandros y redujo 230 Km la longitud del río. Un 13 % de los 266 reservorios (de 2.500 a 12.000 Ha) que existen en la cuenca se localizan sobre el curso del Mississippi (West Consultants, 2000b).
En base a sus características geomorfológicas se reconocen diferentes tramos (Fremling et al., 1989). La alta cuenca se extiende desde las nacientes hasta las cataratas de San Antonio, es un tramo torrentoso y corre por terrenos escarpados. Se contabilizan 11 represas y 6 embalses para control de inundaciones considerados de escaso impacto ambiental. Le sigue el Mississippi Superior (hasta la confluencia con el río Missouri), con la extraordinaria relevancia de atravesar zonas densamente pobladas. Cobra gran importancia como vía de navegación y exhibe condiciones propias de un río con llanura aluvial. Posee 27 embalses escalonados "pools", para mantener el agua dentro de los límites del cauce lleno "bankfull", que modifican fuertemente el paisaje (Figura 2). Los embalses poseen un área léntica que correspondía a la llanura de inundación y otra lótica, hacia la cola del embalse, con presencia de islas, humedales costeros y canales secundarios (West Consultants, 2000a).

Fig. 2. Tramo superior del río Mississippi mostrando en corte la sucesión de embalses (pools) construidos.
Los embalses redujeron las variaciones de nivel hidrométrico, de velocidades de corriente y aumentaron el tiempo de residencia del agua en la llanura aluvial, lo que debilitó las raíces y produjo pérdidas en la vegetación terrestre y también alteró los patrones sedimentológicos, hidrológicos y bióticos, favoreciendo la fragmentación del paisaje.
Muchos de los embalses del Mississippi superior presentan un gradiente ambiental muy marcado que define un ambiente lacustre próximo a la presa y otro lótico hacia la cola de los embalses con extensos humedales. Las obras de dragado, albardones y espigones laterales influyeron para dirigir el caudal hacia el thalweg promoviendo la reducción de los cauces secundarios. A su vez, la construcción de empalizadas y terraplenes modifica el intercambio de agua, sedimentos y materia orgánica con la planicie, disminuyendo la superficie inundada. La Figura 3 presenta un esquema del aspecto que posee actualmente ciertos sectores del Mississippi Superior, la formación de los embalses para navegación, la construcción de espigones para dirigir el flujo, el revestimiento de costas y el uso de la llanura aluvial para agricultura y urbanizaciones.
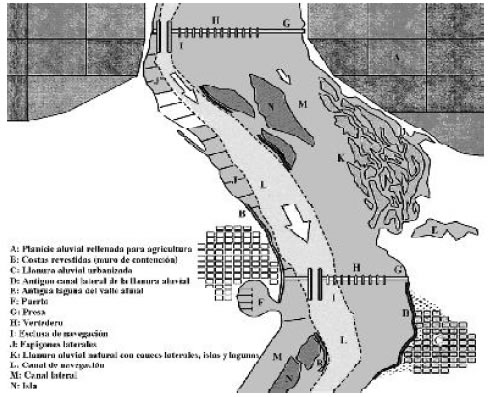
Fig. 3. Esquema de un embalse de navegación ("pool") mostrando las modificaciones ocasionadas por el uso de la planicie aluvial y estructuras construidas en el cauce. La flecha indica el sentido de la corriente y el ancho la velocidad
A su vez la reducción de las llanuras de inundación estimuló el avance de la agricultura sobre los fértiles terrenos de las planicies. Se estima que un 66% del valle de inundación original del Mississippi fue modificado por prácticas agrícolas y que sólamente un 23% de la llanura aluvial mantiene las condiciones naturales. En el Mississippi superior y sus afluentes existe una progresiva y dramática pérdida de las planicies de inundación, siendo reemplazadas de Norte a Sur por tierras cultivadas (Figura 4).
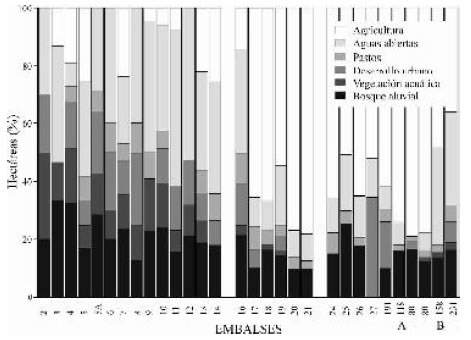
Fig. 4. Superficie de los ambientes que se observan en los embalses (pools) del río Mississippi Superior. A: posición en la milla del río Mississippi. B: posición en la milla del río Illinois.
El Mississippi Medio se extiende entre la confluencia de los ríos Missouri y Ohio y al igual que el tramo anterior, los terraplenes y espigones laterales modificaron los patrones hidrológicos, sedimentológicos y el ingreso de agua a la llanura aluvial, observándose, en el estiaje, desecamiento de cauces secundarios.
Por último el bajo Mississippi corresponde al tramo hasta la desembocadura en el Golfo de México con la llanura de inundación más desarrollada. Sin embargo, es donde mejor se aprecia la interacción con otros ecosistemas que dependen en alguna medida de la cuenca del Mississippi.
La regulación del régimen hidrológico del río a lo largo de todo el curso modificó el patrón de crecientes y bajantes que dejaron de ser marcadamente estacionales para convertirse en irregulares y atenuados, aumentando las crecidas extraordinarias.
Asimismo se observó una reducción dramática en la tasa de transporte de sedimentos, que disminuyó un 75 %, y favoreció la salinización de los humedales costeros en el golfo de México (Martín et al., 2002). Atribuido a los numerosos embalses construidos en toda la cuenca y a la pérdida de las llanuras de inundación (West Consultants, 2000b). Asimismo, aumentó de transporte de nutrientes debido a la actividad agrícola y la pérdida de humedales riparios que actuaban como filtros naturales, generando hipoxia en las aguas costeras (Mitsch et al., 2001).
El Paraná medio: Valor de la complejidad estructural y de los procesos ecológicos en grandes ríos
El río Paraná (Figura 1) tiene una extensión de 2.570 Km y nace de la confluencia de los ríos Paranaíba (de 1.200 Km) y Grande (en Brasil). Tiene una antigüedad de 3 a 4 millones de años (Paoli et al., 2000). Juntamente con los ríos Paraguay (2.550 Km), Uruguay (1.612 Km) y el Río de la Plata (250 Km), conforman la cuenca del Plata con más de 3.1 x 106 Km2 Paoli et al., 2000. Abarca completamente el Paraguay y porciones muy importantes del SW de Brasil, NE de Argentina, SE de Bolivia y SW de Uruguay. En términos del área de drenaje, es el segundo de América del Sur y el quinto en el mundo (Welcomme, 1985). Está dividido en cuatro tramos el superior, alto, medio e inferior (Bonetto, 1994). El Alto Paraná se extiende desde los Saltos del Guairá hasta la confluencia con el río Paraguay y al igual que todos los afluentes del tramo superior están casi completamente represados, con el casi único objetivo de producir electricidad. Hasta la altura de la ciudad de Posadas (Argentina) el cauce principal del río Paraná y todos los afluentes siempre ocuparon el mismo espacio, lo que dio lugar a un cañón. Aguas abajo de Posadas, marca un cambio importante al tomar un carácter algo anastomosado hasta la confluencia con el río Paraguay y no presenta una llanura aluvial definida. Aquí se construyó Yacyretá.
El Paraná Medio se extiende desde la confluencia con el río Paraguay hasta la ciudad de Diamante (Pcia. de Entre Ríos) definiendo una cuenca de 370.000 Km2. Presenta un cauce anastomosado meandriforme; corre por una falla tectónica dejando a su derecha una llanura aluvial de 19.200 Km2, que oscila entre 13 y 60 Km de ancho. El régimen hidrológico del Paraná Medio, si bien está alterado por el conjunto de represas construidas en la alta cuenca igualmente presenta un complejo patrón de crecidas anuales (Paoli y Cacik, 2000). Es posible encontrar un pico principal de crecida en la primavera tardía y el verano, de enorme importancia por sus implicancias ecológicas y un pico secundario en el otoño, debido a la influencia del río Paraguay y ocasionales crecidas provenientes del río Iguazú. La descarga media anual es de 17.000 m 3/s, pero en los últimos 25 años se han observado importantes alteraciones en régimen de crecidas, atribuidos a un cambio en los regímenes de lluvias, deforestación y construcción de represas (Figura 5).
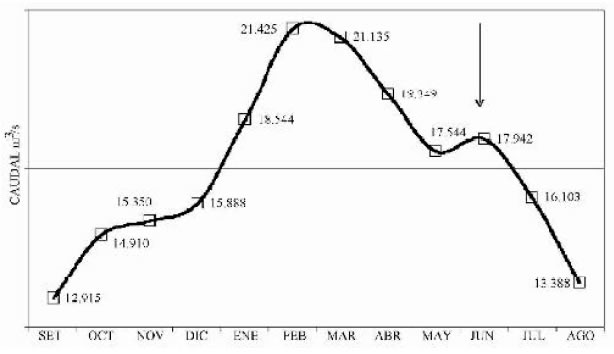
Fig. 5. Caudales medios mensuales cronológicos históricos del río Paraná a la altura de la ciudad de Corrientes. La flecha indica el pico secundario originado por la crecida del río Paraguay.
El Paraná Medio, acarrea una importante carga de sedimentos suspendidos con alto contenido de material fino. El Bermejo aporta entre el 50 y 80% del total (Bonetto, 1994; Bertolino y Depetris, 1992) y pueden alcanzar una concentración de 4.500 mg/l (Drago y Amsler, 1981). La textura del lecho del canal principal está compuesto por arenas medias (Drago y Amsler, 1988). Según Depetris y Lenardon (1982) la tasa media de transporte de sedimentos es de 79,4 millones de toneladas por año y 38,7 millones de toneladas para un período seco.
La llanura aluvial del Paraná, según Drago (1973) es tipo "fringe floddplain" un ecosistema muy complejo compuesto por meandros abandonados, barras, deltas internos, lagunas semicirculares, cauces secundarios, riachos, etc. El ancho varia desde 13 Km a la altura de la confluencia con el río Paraguay hasta casi 60 Km frente a Rosario (Paraná Inferior). Esta planicie aluvial esta cruzada por pequeños cursos donde se alternan sectores playos con pozones profundos de 5 a 15 m (Marchese et al., 2002). La presencia de lagunas someras representa un carácter distintivo de la llanura de inundación con profundidades que no exceden los 5 metros y fondos limo-arcillosos en el centro de sus cubetas.
La dinámica hídrica del valle aluvial está estrechamente dependiente de los pulsos de inundación; condiciona y regula importantes procesos geomorfológicos, biogeoquímicos y la estructura de las comunidades bióticas. La planicie posee islas, islotes, bancos de arena que se desplazan dependiendo del balance erosión-deposición. Las islas poseen madrejones de forma redondeada y lagunas interiores, usualmente alargadas con conexiones semi-permanentes.
La producción en el valle aluvial depende de los pulsos de inundación, que gobiernan la persistencia de la comunidades bióticas (Power et al., 1995) y representan uno de los elementos fundamentales para mantener la integridad ecológica (Poff et al., 1997). El balance de las comunidades, los patrones de biodiversidad y la organización funcional dependerían del efecto anual de la crecida que transporta una gran cantidad de carbono orgánico disuelto alóctono que sufre un procesos de metabolización (Depetris y Paolini, 1991). Esta variación del carbono disuelto en la planicie, se correlaciona con los cambios en el nivel hídrico (Depetris y Cascante, 1985), el contenido de materia orgánica acumulado en los suelos inundados y la tasa de descomposición de la materia orgánica particulada. Moviliza y resuspende los nutrientes contenidos en los sedimentos y aporta a la productividad primaria de algas y macrófitas (Bayley, 1995). A su vez el detritus formado y acumulado, soporta una rica invertebrofauna y representa la base trófica de varias especies de peces. Al iniciarse la estación seca, importantes cantidades de este carbono disuelto y particulado son exportados al cauce principal.
Los pulsos periódicos de inundación favorecen el ingreso de sólidos en suspensión y el acarreo de nutrientes adsorbidos (Bonetto et al., 1989), disparando el desarrollo del fito y zooplancton (Bonetto, 1975) esencial para la supervivencia de larvas y juveniles de peces en general. El aumento de materia orgánica como detritus constituye la principal fuente de alimentación para juveniles y adultos del sábalo (Bowen, 1983; Fugi et al., 1996) que representaba mas del 50 % de la biomasa total con valores promedio que podían superar los 1.000 kg ha-1 (Bonetto et al., 1970; Bonetto et al., 1969; Bonetto, 1986; Quirós y Baigún, 1985). A su vez, los huevos, larvas y juveniles de sábalo, considerada la especie clave del sistema, constituyen la principal fuente de alimentación de larvas y juveniles de los ictiófagos y predadores topes del sistema (surubíes, dorados y bagres en general) estableciendo una de las principales vías de energía del sistema (Oldani, 1990).
La presencia de especies detritívoras aparece como una característica saliente en grandes ríos latinoamericanos con llanura de inundación (Bayley, 1991; Welcomme, 1985). La fauna bentónica, a su vez, presenta una respuesta directa a la dinámica de inundación, variando la composición a lo largo de un gradiente definido por la conexión entre el canal principal, los cauces secundarios y las lagunas de la planicie (Marchese et al., 2002).
El desarrollo de la vegetación acuática asociado a las variaciones del nivel hidrométrico juega un papel fundamental en el ciclo de la materia orgánica y nutrientes (Neiff, 1978). Las macrófitas flotantes y arraigadas alteran la calidad del agua, favorecen la decantación de los sedimentos y proporcionan un hábitat adecuado para refugio y cría de peces y el desarrollo de la importante comunidad de invertebrados acuáticos (Poi de Neiff et al., 1994).
Discusión
Los grandes ríos con llanura de inundación representan sistemas complejos de importante valor ecológico. Proveen áreas de refugio, alimentación, reproducción y cría para diversas especies y actúan como corredores de biodiversidad para las especies migradoras (Sparks, 1995; Welcomme, 1985). Exhiben ciclos biogeoquímicos complejos donde las comunidades biológicas están indisolublemente ligadas a los cambios de las características ambientales. Diferentes paradigmas trataron de reflejar estos aspectos. El concepto pionero del "continuum" introducido por Vannote et al., (1980), puso en evidencia la importancia de los procesos de transporte longitudinal de materia orgánica y nutrientes y su relación con el aprovechamiento por parte de la biota. Un aspecto central relacionado con este concepto son los desplazamientos y el reciclado de los nutrientes y de la materia orgánica que se produce unidireccionalmente aguas abajo. Este fenómeno conocido como espiralado de nutrientes (Elwood et al., 1983) se vincula con la dinámica energética del sistema debido a la interacción entre el transporte de nutrientes, el arrastre de carbono y los procesos biológicos y ecológicos que derivan de la oxidación de carbono orgánico. Por otra parte, el concepto de pulso de inundación desarrollado por Junk et al., (1989) emergió como un modelo conceptual apropiado para grandes ríos con llanura aluvial. Refleja adecuadamente el intercambio de materia y energía que se produce entre el cauce principal y la llanura de inundación gracias a la recurrencia de los pulsos. Este último enfoque pone de manifiesto la importancia de una libre conectividad entre los subsistemas, siendo el elemento crítico para mantener la estructura de la llanura y preservar los procesos que regulan la producción del río a través del intercambio de nutrientes y materia orgánica. Estos procesos se interrumpen cuando los ríos son regulados con represas debido a que se modifican los factores bióticos y abióticos, lo que se refleja en el concepto de discontinuidad seriada (Ward y Stanford, 1983). Claramente la producción biótica es mayor en aquellos ríos con planicies libremente conectadas. Bayley (1995), presentó evidencias de que las lagunas aisladas del canal principal del Mississippi tenían un 64% menos de peces que las que mantenían una conexión periódica, mientras en el río Missouri una reducción del 67% del área inundada de la llanura aluvial, disminuyó un 80% las capturas de peces (Whitley y Campbell, 1974).
Estos paradigmas se cumplen en ríos que no han sido sometidos a disturbios pronunciados y reflejan los procesos que se manifiestan para mantener la integridad ecológica de estos sistemas. Por esto, los intentos de rehabilitación y particularmente de restauración, deben estar dirigidos a recuperar dichos procesos. De acuerdo a Lewis et al., (2000) la restauración de los grandes ríos debería comenzar por recobrar el funcionamiento de las llanuras aluviales, que son áreas de transición acuático-terrestre (Junk et al., 1989) y poseen características ecológicas únicas. Actúan como ecotonos donde se verifican procesos hidro-geomorfológicos que definen un rico mosaico de parches ecológicos (Petts, 1990) y la biota está adaptada a explotar temporal y espacialmente la heterogeneidad ambiental (Ward et al., 1999). La presencia de lagunas, albardones, bancos de arenas, cauces secundarios y meandros les confiere una compleja estructura, mientras que los pulsos de inundación representan el motor que viabiliza los ciclos biogeoquímicos fundamentales.
Sparks (1995) concluye que la restauración de las planicies de inundación representa un paso obligado para recuperar la capacidad de intercambiar materia orgánica con el río que se encuentra directamente ligado a la capacidad de disparar procesos claves como el flujo de materia orgánica y el metabolismo del carbono. Estos procesos se encuentran severamente alterados en el Mississippi, siendo las condiciones de flujos de sedimentos y carbono muy diferentes de los originales. El avance sostenido en el deterioro de los grandes ríos como: Mississippi, Danubio y Rhin, etc. llevo a plantear mecanismos adecuados para recomponer la integridad ecológica y revertir las trayectorias con propiedades ecológicas visiblemente modificadas que conducen a una virtual extinción de los sistemas.
Si bien se desarrollaron numerosos estudios sobre restauración de ríos de bajo orden, existe la percepción de que los resultados difícilmente puedan ser extrapolados a grandes ríos (Sparks, 1995) con escalas temporal-espacial muy diferentes. La Tabla 1 compara características estructurales y funcionales entre ríos de bajo orden y grandes ríos con llanuras de inundación y confirma la necesidad de establecer criterios diferentes de restauración.
Los grandes ríos representan en general sistemas con alto nivel de impacto en sus cuencas (Gore y Shields, 1995). Esta situación no se aplica aún a los tramos medios e inferior del Paraná los que ofrecen la posibilidad de estudiar un ecosistema fluvial y aluvial donde ocurren procesos naturales que regulan el funcionamiento como ecosistemas integrados. Estos sectores poseen un valle de inundación escasamente perturbado, donde se verifica una libre conectividad entre los diferentes componentes (cauces principales y secundarios, planicies, lagunas interiores, etc.). Esto contrasta con el Mississippi, que plantea diferentes escenarios de acuerdo a las políticas de manejo y gestión que se desarrollen en una u otra cuenca.
En el Mississippi se adoptaron diferentes estrategias de restauración centradas en la rehabilitación de algunos tramos, donde el mayor obstáculo probablemente sea la imposibilidad de recuperar la perspectiva ecológica histórica de la cuenca, modificada desde hace más de 100 años y con escasos sectores donde aún se observan condiciones de baja perturbación. Galat et al., (1998) señalan que únicamente bajo condiciones extraordinarias como lo ocurrido en la inundación de 1993, es posible aproximarse a un escenario que refleje el comportamiento de la biota como respuesta al pulso de inundación. En este contexto, Sparks et al., (1990) notan que la comprensión que los disturbios antrópicos que se originan al regularse los grandes ríos, requieren necesariamente de una comparación con sistemas no alterados.
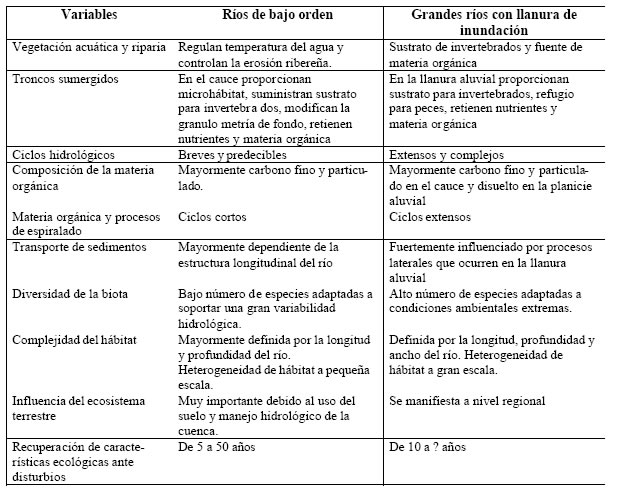
Tabla 1. Comparación entre procesos funcionales y características estructurales en ríos de bajo orden y grandes ríos con llanura de inundación.
El Paraná medio soporta impactos que pueden ser considerados moderados. Las perturbaciones más importantes son, una ruta que atraviesa todo el valle aluvial entre las ciudades de Santa Fe y Paraná y la construcción de algunos polders en áreas próximas a las ciudades. También se registra sobreexplotación pesquera, contaminación de agroquímicos, deforestación, uso ganadero de las islas, dragado, contaminación con metales pesados provenientes sobre todo de explotaciones mineras en Bolivia. El régimen hidrológico del río ha sido alterado en parte por el cambio climático global y el manejo de las represas en la alta cuenca y exhibe una llanura de inundación que no fue modificada. Estos impactos moderados pueden ser revertidos y devolver el sistema a una condición próxima a la original, simplemente basándose en su capacidad de resiliencia. En cambio en el Mississippi, esta situación ya no es posible debido a que posee un nuevo estado de equilibrio (inestable) donde las condiciones ecológicas que rigen el comportamiento del sistema fueron dramáticamente modificadas con: pérdidas de llanura de inundación, atenuación de pulsos de crecida, entrampamiento de sedimentos, bloqueo de las migraciones de peces, reducción de intercambio de sedimentos entre el cauce y la llanura, etc. Aspectos que impiden virtualmente rehabilitar el Mississippi, debiéndose poner énfasis en preservar completamente las áreas que aún exhiben un estado natural adecuado y restaurar aquellos sectores que presentan bajos impactos. Esta estrategia permitiría llevar al río a recuperar parcialmente la integridad ecológica perdida.
Claramente las condiciones presentes en el Paraná medio representan una razonable aproximación a la situación en la que se encontraba la cuenca del Mississippi 200 años atrás y como se han ido modificando sus características de acuerdo al aumento de impactos, tales como represamientos, uso de la planicie de inundación, construcción de estructuras para regular el caudal, etc. (Figura 6). Sin duda, el Paraná puede proporcionar importantes claves para encarar la rehabilitación, restauración y remediación de otros ríos, pero la historia del Mississippi representa también un espejo y un serio llamado de atención para mirar un posible escenario de los ríos que aún mantienen cierto grado de condiciones naturales. La cadena de embalses y represas que hoy cortan el Mississippi en su tramo superior, por ejemplo, no difieren mayormente en sus consecuencias de los proyectos de represas pergeñados para los ríos Paraná, Uruguay y Paraguay y ya efectivizados en la cuenca superior de los ríos Paraná y Uruguay. Todos, de alguna manera, alteran los ciclos biogeoquímicos al reducir o eliminar las planicies aluviales, generando cambios en su estructura y funciones.
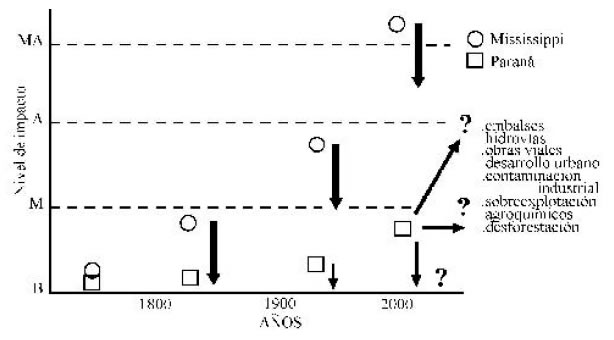
Fig.6. Evolución temporal comparada de los impactos en los ríos Paraná y Mississippi. B: bajo; M: moderado; A: alto; MA: muy alto. La flechas indican el nivel de recuperación que puede alcanzarse aplicando técnicas de rehabilitación y /o restauración según el caso.
La pérdida de la integridad ecológica de los grandes ríos implica asimismo pérdidas económicas importantes para muchos sectores que utilizan los servicios de estos ecosistemas. La construcción de embalses en la cuenca superior (Brasil), por ejemplo, generó una caída importante en los rendimientos pesqueros (Agostinho et al., 1994; Okada et al., 1996) y en la calidad de las pesquerías (Agostinho et al., 1995).
Este proceso puede estar ocurriendo en Yacyretá, mientras que aguas arriba en el Alto Paraná (Posadas-Puerto Iguazú), las pesquerías comerciales virtualmente desparecieron por la interrupción de las migraciones de especies de alto valor económico (Baigún, com. Pers.). Lo anterior señala la conveniencia de estimar los costos de recuperación de los ríos con las pérdidas devengadas de su degradación.
Se concluye que el ejemplo de estos dos sistemas en situaciones casi opuestas, permite ejemplificar fases del proceso de transformación que sufrieron los grandes ríos con llanuras de inundación. La falta de percepción adecuada sobre la necesidad de no alterar severamente los procesos fundamentales que sustentan la integridad ecológica de estos sistemas, condujeron al Mississippi a un estado de degradación, que no puede ser revertido a su condición original. Ésta es la lección fundamental que debe aprenderse para evitar un destino similar para el Paraná y su cuenca. Sólamente un manejo racional y conservacionista de los diferentes ecosistemas que componen su cuenca baja y media, incluyendo a la del río Paraguay, permitirá evitar un deterioro similar al del Mississippi y otros grandes ríos, y torcer la historia hacia una trayectoria divergente.
Bibliografía
Agostinho, A. A., H. F. Julio y M. Petrere Jr. 1994. Itaipú reservoir (Brasil): Impacts of the impoundment on the fish fauna and fisheries. En: Cowx, I. G. (ed.) Rehabilitation of freshwater fisheries. Fishing New Books, Oxford, United Kingdom: 171-184.
Agostinho, A. E. Vazzoler y S. M. Thomaz. 1995. The high Paraná basin: limnological and icthyological aspects. En: Tundisi, J. G., C. E. Bicudo y T. Matsamura Tundisi (eds.) Limnology in Brazil. Brazilian Academy of Sciences: 60-103.
Bayley, P. B. 1991. Fish yield from the Amazon from the Amazon in Brazil: comparisons with African river yields and management possibilities. Transactions of the American Fisheries Society. 110: 351-359.
Bayley, P. B. 1995. Understanding large river-floodplain ecosystems. BioScience 45: 153-158.
Bertolino, S. R. y P. J. Depetris. 1992. Mineralogy of the clay–sized suspended load from headwater tributaries on the Paraná River: Bermejo, Pilcomayo, and Paraguay Rivers. En: E. T. Degens, S. Kempe, A. Lein and Y. Sorokin (eds.) Interactions of biogeochemical cycles in aqueous ecosystem. Pt. 7. Mitteilungen des Geologish-Paläeontologischen Instituts der Universität Hamburg, SCOPE/UNEP Sonderband 52: 19-31.
Bonetto, A. A. 1975. Hydrologic regime of the Paraná River and its influence on ecosystems. En: Hastler (ed.) Coupling of land and water systems, Springer Verlag, New York: 175-197.
Bonetto, A. A., 1986. Fish of the Paraná System, The ecology of River System, Davies, B. R. y K. F. Walker (eds.), Dordrecht, The Netherlands, 574 p.
Bonetto, A. A. 1994. Austral rivers of South America. En: R. Margalef (ed.) Limnology Now, Elsevier Science B.V, Amsterdam, The Netherlands: 425-472.
Bonetto, A. A., E. Cordiviola de Yuan, C. Pignalberi y O. Oliveros, 1969. Ciclos hidrológicos del río Paraná y las poblaciones de peces contenidas en las cuencas temporarias de su valle de inundación. Physis 29: 213-223.
Bonetto, A. A., E. Cordiviola de Yuan, C. Pignalberi y O. Oliveros 1970. Nuevos datos sobre poblaciones de peces en ambientes permanentes del Paraná Medio. Physis 30: 141-154
Bonetto, A. A., I. Waiss y H. Castello. 1989. The increasing damming of the Paraná basin and its effects on the lower reaches. Regulated Rivers 4: 333-346.
Bowen, S. H. 1983. Detritivory in Neotropical fish communities. Environmental Biology of Fishes 9: 137-144.
Depetris, P. J. y A. M. Lenardón. 1982. A second report on particulate and dissolved phases in the Paraná River. En: Degens, E. T, S. Kempe y H. Soliman (eds.) Transport of carbon and minerals in major world rivers. Pt 2. Mitteilungen des Geologish-Paläeontologischen Instituts der Universität Hamburg, SCOPE/UNEP Sonderband 52: 167-181.
Depetris, P. J y E. Cascante. 1985. Carbon transport in the Parana River. En: Degens, E. T, S. Kempe y R. Herrera (eds). Transport of carbon and minerals in major world rivers. Mitteilungen des Geologish-Paläeontologischen Instituts der Universität Hamburg, SCOPE/UNEP Sonderband 52: 385-395.
Depetris, P. J. y J. E. Paolini. 1991. Biogeochemical aspects of South American Rivers: The Paraná and the Orinoco. En: Pt. 1. Mitteilungen des Geologish-Paläeontologischen Instituts der Universität Hamburg, SCOPE/UNEP Nro 42, John Wiley, New York:105-125.
Drago, E. 1973. Caracterización de la llanura aluvial del Paraná Medio y de sus cuerpos de agua. Boletín Paranaense de Geociencias 31: 31-44
Drago, E. C. y M. L. Amsler. 1981. Sedimentos suspendidos en el tramo medio del río Paraná: variaciones temporales e influencia de los principales tributaries. Revista de la Asociación de Ciencias Naturales del Litoral 12: 28-43.
Drago, E. C. y M. L. Amsler. 1998. Bed sediment characteristics in the Paraná and Paraguay Rivers. Water International 23: 174-183.
Elwood, J.W., J. D. Newbold, R. V. O‘Neill y W. van Winkle. 1983. Resource spiraling: an operational paradigm for analyzing lotic ecosystems. En: Fontaine, T. D y S. M. Bartell (eds.) Dynamics of lotics ecosystems. Ann Arbor Science: 3-28.
Fremling, C. R., J. L. Rasmussen, R. E. Sparks, S. P. Cobb, C. F. Bryan and T. O. Clafin. 1989. Mississippi River fisheries: a case history. Proceedings of the international large river symposium (D. P. Dodge, ed). Canadian Special Publication in Fisheries and Aquatic Sciences 106: 309-351.
Fugi, R. N. S. Han y S. Agostinho. 1996.. Feeding strategies of five species of bottom-feeding fish of the high Parana River (PR-MS). Environmental Biology of Fishes 46: 297-307.
Galat, D. L., H. L. Fredrickson, D. Humburg, K. Bataille, J. Russell Bodie, et al. 1998. Flooding to restore connectivity of regulated, large-river wetlands. BioScience 48: 721-733.
Gore, J. A. y F. D. Shields. Jr. 1995. Can large rivers be restored?. BioScience 45: 142-152.
Junk, W. J., Bayley, P. B. y Sparks, R. E. 1989. The flood pulse concept in river-floodplain systems. En: Dodge, (D. P, ed.) Proceedings of the International Large River Symposium. Canadian Special Publication of the Fisheries and Aquatic Science, 106: 110-127.
Lewis, W. M. S. K. Hamilton, M. A. Lasi, M. Rodríguez y J. Saunders. 2000. Ecological determinism on the Orinoco floodplain. BioScience 50: 681-592.
Marchese, M., I. Ezcurra de Drago y E. C. Drago. 2002. Benthic macroinvertebrates and ohysical habitat relationships in the Paraná river flood-plain system. En: M. E. McClain (ed) The ecohydrology of South American rivers and wetlands. IAHS Special Publication 6: 111-132.
Martin, J. E., E. Reyes, G. P. Kemp, H. Mashriqui y J. Day Jr. 2002. Landscape modeling of the Mississippi Delta. BioScience 52: 357-365.
Mitsch, W. J., J. W. Day Jr., J. W. Gilliam, P. M. Groffman, D. H. Hey, G. W. Randall y N. Wang. 2001. Reducing nitrogen loading to the Gulf of Mexico from the Mississippi River basin: strategies to counter a persistent ecological problem. BioScience 51: 373-388.
Neiff, J. J. 1978. Fluctuaciones de la vegetación acuática en lagunas del valle del río Paraná en la transección Paraná-Santa Fe entre 1970 y 1977. Physis 38 (Buenos Aires): 41-53.
Okada, E. K., A. A. Agostinho y M. Petrere Jr. 1996. Catch and effort data and the management of the commercial fisheries of Itaipú reservoir in the upper Parana River, Brazil. En: Cowx, I. (ed.) Stock assessment in inland water fisheries. Fishing New Books, London, United Kingdom: 164-171.
Oldani, N. 1990. Variaciones de la abundancia de peces del valle del río Paraná (Argentina). Revue d´Hydrobiologie Tropicale 23: 67-78.
Paoli, C; M. Iriondo y N. García 2000. Características de las cuencas de aporte. En: Paoli, C. y M. Schreider (eds.) El río Paraná en su tramo medio. Contribución al conocimiento y prácticas ingenieriles en un gran río de llanura. Universidad Nacional del Litoral, Santa Fe: 30-68.
Paoli, C. y P. Cacik. 2000. Régimen de crecidas y análisis de caudales máximos. En: Paoli, C. y M, Schreider (eds.) El río Paraná en su tramo medio: contribución al conocimiento y prácticas ingenieriles en un gran río de llanura. Universidad Nacional del Litoral, Santa Fe: 109-171.
Petts, G. E. 1990. The role of ecotones in aquatic landscapes management. En B. Neiman y H. Decamps (eds) The roles of ecotones in aquatic landscapes. Parthenon Press, London Press: 227-261.
Poff, N. L., J. D. Allan, M. B. Bain, J. R. Karr, K. I. Prestegaard, B. D. Richter, R. E Sparks y J. C. Stromberg. 1997. The natural flow regime: a paradigm for river conservation and restoration. BioScience 47: 769-784.
Poi de Neiff, A., J.J Neiff, O. Orfeo y R. Carignan. 1994. Quantitative importance of particulate matter retention by the roots of Eichhornia crassipes in the Paraná floodplain. Aquatic Botany 47: 213-223.
Power, M. E., A. Sun, G. Parker, W. E. Dietrich y J. Timothy Wooton.1995. Hydraulic food-chain models. BioScience 45: 159-167.
Quirós, R. y C. Baigún. 1985. Fish abundance related to organic matter in the Plata River Basin, South America. Transactions of the American Fisheries Society 114: 377-387.
Sparks, R. K. 1995. Need for ecosystem management of large rivers and their floodplain. BioScience 45: 168-182.
Sparks, R. E., P. B. Bayley, S. L. Kohler y L. L. Osborne. 1990. Disturbance and recovery of large floodplain rivers. Environmental Management 14: 699-709.
Vannote, R.L., G. W. Minshall, K. W. Cummins, J. R. Sedell y C. E. Cushing. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37: 130-137.
Ward, J. V. y J. A. Stanford. 1983. The serial discontinuity concept of lotics ecosystems. En: Fontaine, T. D y S. M. Bartell (eds.) Dynamics of lotics ecosystems. Ann Arbor Science: 29-42.
Ward, J. V., K. Tockner y F. Schiemer. 1999. Biodiversity of floodplain river ecosystems: ecotones and connectivity. Regulated Rivers Research and Management 15: 125-139.
Welcomme, R. L. 1985. River Fisheries, FAO Fisheries Technical Paper, No. 262, Food and Agriculture Organization of the United Nations.
West Consultant. 2000a.Upper Mississippi River and Illinois waterways cumulative effects study. Volume II: Ecological assessment. Environmental Report 40-2. Contract NO.DACW 25-97-R-0012.
West Consultant. 2000b.Upper Mississippi River and Illinois waterways cumulative effects study. Volume I: Geomorphic assessment. Environmental Report 40-1. Contract NO.DACW 25-97-R-0012.
Whitley, J. R. y R. S. Campbell. 1974. Some aspects of water quality and biology of the Missouri River. Transactions of Missouri Academy of Science. 8: 60-72.