Peces marinos y continentales del Mioceno del área mesopotámica
argentina. Edad y relaciones biogeográficas.
Alberto Luis CIONE1
; Jorge Rafael CASCIOTTA2
; María de las Mercedes AZPELICUETA2
; María Julia BARLA3
y Mario Alberto COZZUOL4
1Division Paleontología de Vertebrados.Facultad de Ciencias Naturales y Museo. 1900, La Plata.
E-mail: acione@museo.fcnym.unlp.edu.ar
2Division Zoología Vertebrados. Idem. E-mail: azpeli@museo.fcnym.unlp.edu.ar,
jrcas@museo.fcnym.unlp.edu.ar
3Facultad de Ciencias Naturales y Museo. 1900, La Plata. E-mail: barla@netverk.com.ar
4Laboratorio de Biología Evolutiva e da Conservaçao Universidade de Rondonia. BR 364, Km 9,5. 78900-000
Porto Velho, Ro, Brasil. E-mail: mario@unir.br
Abstract: MARINE AND CONTINENTAL FISHES OF MIOCENE IN THE ARGENTINE MESOPOTAMIAN AREA.AGE AND BIOGEOGRAPHIC RELATIONS.- Recently, a new locality with brazilian fishes was described from the Miocene of Patagonia by Cione and others. However, the most renown localities are located in the Paraná eastern riverside cliffs near the city of Paraná, Entre Ríos province, Argentina. These localities yielded many freshwater, marine, and terrestrial vertebrates of Miocene and Pleistocene age which in many cases are presently in study. Miocene fossils come almost exclusively from the Paraná Formation (marine and freshwater taxa: elasmobranchs, teleosteans, cetaceans, sirenians and pinnipeds) and the “Conglomerado osífero” (“Bony conglomerate;” “Mesopotamiense” autorum) at the base of the continental Ituzaingó Formation (marine, freshwater and terrestrial taxa: elasmobranchs, teleosteans, crocodilians, chelonians, birds, and different groups of mammals).
Cetaceans suggest a Tortonian (late Miocene) age for the top of the Paraná Formation. The terrestrial mammal fauna suggests a Huayquerian age (Tortonian) for the base of the Ituzaingó Formation. Both the terrestrial and freshwater fauna of the Ituzaingó Formation indicates a climate warmer than present. The freshwater vertebrate record suggests important basin conections with Amazonian basins.
Key words: Fishes - Miocene - Entre Ríos - Tertiary
Palabras clave: Peces - Mioceno - Entre Ríos - Terciario.
Introducción
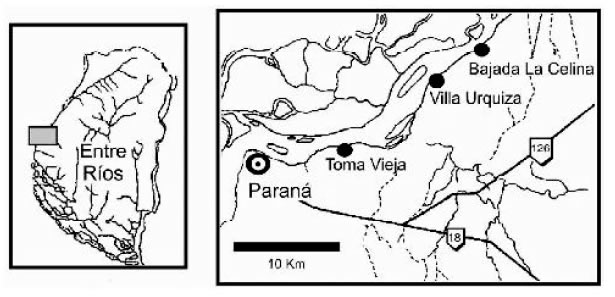
Los yacimientos fosilíferos de la barranca oriental del río Paraná cercanos a la ciudad de Paraná (este de Argentina) se conocen desde 1827 cuando Alcide D’Orbigny visitó el área (véase D’Orbigny, 1842; Figura 1). Las barrancas son extremadamente ricas en restos de vertebrados terrestres, marinos y de aguas continentales. Sin embargo, la mayoría de las colecciones se han hecho sin una adecuada ubicación estatigráfica. Debido a ésto, durante muchos años se debatió la procedencia de los fósiles (Cione et al., 2000). Trabajo de campo reciente realizado por los autores han permitido clarificar la ubicación estratigráfica de los vertebrados. Este trabajo ha permitido colectar materiales de vertebrados marinos en la Formación Paraná en Aldea Brasilera (cerca de Diamante) y Villa Urquiza y vertebrados acuático continentales y terrestres en la Formación Ituzaingó en la Toma Vieja y La Celina (ambas localidades cerca de la ciudad de Paraná (Figura 1).
En esta contribución, se analiza el registro de peces continentales y marinos de las formaciones Paraná e Ituzaingó en las cercanías de Paraná y se analiza preliminarmente su composición. Ciertamente, las investigaciones se encuentran en progreso y probablemente muchas de las asignaciones taxonómicas serán modificadas en los próximos años y nuevos taxones serán incluidos en las listas debido a nuevos hallazgos o al estudio de colecciones previamente estudiadas.
Fig. 1. Mapa de ubicación.
Historia del conocimiento de los peces miocenos de Paraná
Cuando Florentino Ameghino, en su importante obra sobre los mamíferos extintos de Argentina (1889), discutió los vertebrados del «Piso Mesopotámico» de Doering (1882), incluyó alrededor de 90 especies de mamíferos además de los peces y reptiles. Este trabajo de Ameghino fue sumamente importante para el conocimiento de los vertebrados del Mioceno del área, el que comenzó con la visita del investigador francés Alcide D´Orbigny en 1827. D´Orbigny llegó a América en 1826; en los siguientes años exploró la barranca del río Paraná cercana a la ciudad the «Bajada de Santa Fé,» como fue llamada al principio la ciudad de Paraná y más tarde (1842) describió la estatigrafía y también muchos fósiles.
Otro famoso científico, Charles Darwin, visitó el área durante octubre del año 1833. Darwin describió la estatigrafía de las barrancas cerca de Paraná. En los sedimentos cercanos a la base de las barrancas fluviales colectó dientes de tiburones e invertebrados de especies extintas. Por encima de este nivel, Darwin encontró un yacimiento consolidado que pasaba por encima de sedimentos pampeanos típicos. Darwin interpretó ésto como el resultado de una gran ingresión marina la cual fue gradualmente transformada en un estuario fangoso donde los cadáveres habrían flotado (Darwin, 1839).
El General Justo José de Urquiza, primer presidente de la Confederación Argentina, con capital en la ciudad de Paraná, propició el comienzo de estudios sistemáticos, geológicos y paleontológicos en la barranca del río. Urquiza contrató entre otros al belga Alfred Du Gratty, quien en 1854 fue el director del Museo de la Confederación en Paraná, al físico y naturalista francés Martin de Moussy, quien estuvo a cargo del relevamiento del territorio nacional; y al arquitecto francés (y paleontólogo aficionado), August Bravard, quien fue el primero en investigar depósitos minerales.
Bravard fue nombrado en 1857 director del Museo de la Confederación. Un año más tarde el Registro Oficial de la Confederación publicó la «Monografía de los terrenos marinos terciarios de las cercanías de Paraná» escrito por Bravard. Este trabajo fue la recopilación de sus investigaciones previamente comunicadas en las sociedades de historia natural de Buenos Aires y París en 1855 y 1856 respectivamente. El trabajo de Bravard concuerda en algunos aspectos con las observaciones de D´Orbigny y Darwin pero presentaba contradicciones con las conclusiones de la obra de De Moussy (1857).
Bravard (1858) describió por primera vez algunos teleósteos y eslasmobranquios, reptiles (incluyendo quelonios, ofidios y cocodrilos) y mamíferos. El presidente Domingo Faustino Sarmiento envió los materiales colectados por Bravard al Museo Nacional de Historia Natural of Buenos Aires. Los mamíferos estaban representados por una ballena y un roedor mas pequeño que «Megamys patagoniensis,» descriptos por D´Orbigny (1842) y procedentes de las barrancas del río Negro cerca de Carmen de Patagones (norte de Patagonia). Bravard consideró que las capas de Paraná y Patagones eran de una edad similar. No estaba muy errado.
En 1861, Bravard murió durante un terremoto que destruyó la ciudad de Mendoza (oeste de Argentina). Después de Bravard (1858), la única publicación en muchos años fue la «Description Physique, Géographique et Statistique de la Confederation Argentine» (De Moussy, 1860).
El Museo de la Confederación fue refundado como el Museo de la Provincia de Entre Ríos en 1884 y el profesor Pedro Scalabrini fue su director desde 1886. Scalabrini fue un gran colector de fósiles y rápidamente se puso en contacto con el gran paleontólogo argentino Florentino Ameghino quien describiría este nuevo material. Esta asociación fue el origen del conocimiento de la fauna del«Piso Mesopotamiense» (Ameghino, 1883a, 1883b, 1885, 1886, 1889).
Entre 1890 y 1892, el suizo Kaspar Jacob Roth (Santiago Roth en la literatura argentina; ver Bond, 1999), relevó las provincias de Entre Ríos y Corrientes. Roth hizo una importante colección de vertebrados del «Conglomerado osífero» de la Formación Ituzaingó, parte de la cual fue depositado en el Museo de La Plata, donde ocupó la jefatura de la Sección Paleontología en 1895.
Durante la última parte de los siglos XIX y comienzo del XX, muchos aficionados colectaron cerca de de Paraná. Más tarde la mayoría de esas colecciones privadas fueron depositadas en el Museo Nacional de Historia Natural de Buenos Aires (colecciones de Lelong Thévenet, de Carles, Sors Cirera y Caixo) y Museo de La Plata (colección de Sors Cirera). Otras instituciones donde se pueden encontrar colecciones del área de Paraná son: Muséum National d´Histoire Naturelle de Paris y Natural History Museum de Londres (originalmente British Museum, Natural History).
Durante el siglo XX, a partir de los importantes trabajos de Frenguelli (1920), muchos autores publicaron contribuciones sobre la paleontología y la geología del área de Paraná (Bianchini y Bianchini, 1971; Aceñolaza, 1976; Aceñolaza y Sayago, 1980; Iriondo y Rodríguez, 1973; Noriega, 1995; Cione et al., 2000; véase la bibliografía citada en este último trabajo).
Comentarios sobre la geología del Cenozoico superior en el área
El mar penetró profundamente en el Mioceno medio-tardío por la llanura chacopampeana («Mid Transgressive Onlap Sequence»; véase Uliana y Biddle, 1988; del Río, 1991; Marengo 2000; del Río et al., 2001). Sin embargo, no parece haber atravesado América del Sur como han sostenido algunos autores del pasado y presente (véase discusión en Aceñolaza, 2000). Esta ingresión fue denominada “Paranense” o “Entrerriense”. En cambio, durante el Oligoceno tardío?-Mioceno temprano el mar había ocupado gran parte de Patagonia sin afectar el área chacopampeana. Esta última fue denominada “Patagoniense” partim o “Leonense”. La ingresión “Entrerriense” sólo afectó muy parcialmente a Patagonia, principalmente en el noreste de la misma. En esa zona, el “Patagoniense” y el “Entrerriense” están separados por una neta discordancia (Feruglio, 1949). Por ejemplo, en el área de Península Valdés y curso inferior del río Chubut, entre las Formaciones Gaiman y Puerto Madryn (Cione, 1988; Cione et al., 2005). Ninguna otra ingresión afectó profundamente a Patagonia posteriormente, lo que es vinculado a la fase de ascenso de la cordillera austral del Mioceno medio (Yrigoyen, 1975).
En el área mesopotámica, la ingresión “Entrerriense” depositó los sedimentos de la Formación Paraná (por ejemplo, Ameghino, 1906; Frenguelli, 1920; Scartascini, 1954; Aceñolaza, 1976; Aceñolaza y Aceñolaza, 1999; Aceñolaza, 2000). En el área no se ha detectado la ingresión “Patagoniense”, la cual estuvo restringida al territorio de Patagonia. La Formación Paraná está principalmente compuesta por limolitas verdosas y areniscas con bancos de ostras (Aceñolaza, 1976; Chebli et al., 1989). Cuando el mar se retiró se formó una red hidrográfica que depositó en los valles sedimentos que son actualmente asignados a la Formación Ituzaingó. Esta unidad está compuesta por un conglomerado basal («Conglomerado osífero») con abundantes restos de vertebrados y que es cubierto por areniscas blancuzcas a amarillentas y limolitas verdes con escasos fósiles. Sobre la Formación Ituzaingó se depositaron sedimentos terrígenos pleistocenos (Aceñolaza, 1976; Iriondo, 1980; Chebli et al., 1989) quizás ensenadenses a ciertamente lujanenses de acuerdo a la escala temporal sudamericana (véase Cione y Tonni, 1995, 1996). Aparentemente la ingresión “Entrerriense” se desarrolló con una marcada influencia de aguas dulces y sólo hay evidencias de ambientes marinos normales en la parte sudoriental (del Río, 1988, 1990; Aceñolaza, 2000; Marengo, 2000).
El término «Piso Mesopotamiense» o «Mesopotamiense» fue ampliamente usado en la literatura argentina de palentología de vertebrados. El término fue introducido por primera vez por Doering (1882; véase también Ameghino, 1883). Frenguelli (1920) restringió el «Mesopotamiense» al “Conglomerado osífero». Nosotros consideramos que es inválido y se utiliza aquí el término “Conglomerado osífero» para denominar el conglomerado que aparece frecuentemente en la base de la unidad litoestratigráfica Formación Ituzaingó.
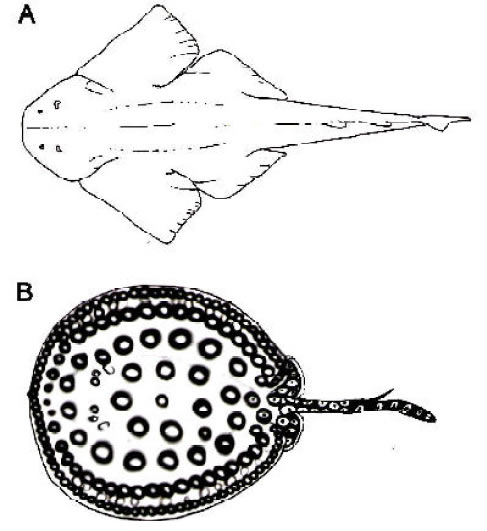
Fig 2. Representantes vivientes de géneros de neoseláceos que se registran en las Formaciones Paraná e Ituzaingó. A Squatina dureril (modificado de Bigelow y Schroeder, 1948). B, Potamotrygon motoro (del natural). No están a escala.
Edad de la Formación Paraná
La edad de la Formación Puerto Madryn ha sido debatida con cierta intensidad (véase del Río
1988, 2000). En años recientes, fue referida al Mioceno medio (del Río 1988, 1990) y más recientemente
al Mioceno tardío (del Río et al. 2001; Scasso et al. 2001). La sección tipo de la correlativa
Formación Barranca Final de Río Negro (Patagonia boreal) fue asignada al Mioceno medio a
tardío(Guler et al. 2002). Concentrados de una capa de ceniza ubicada en los niveles superiores de la
Formación Puerto Madryn en Bahía Cracker, Chubut nororiental, fueron datadas por medio de
40K/39Ar (tres fechados: promedio 9.41 Ma; Zinsmeister et al., 1981). Scasso et al. (2001)
correlacionaron ese nivel con los niveles superiores de la unidad en Puerto Pirámide y en la Lobería
de Puerto Pirámide, en secciones de la Formación Puerto Madryn que estudiaron. Recientemente,
nuevas dataciones de la Formación Puerto Madryn fueron obtenidas por medio de análisis de
87Sr/86Sr en conchas de pectínidos y ostras (Scasso et al. 2001). Los fechados van de 18.4 a 8.7 Ma; los
autores descartaron los fechados más antiguos y retuvieron como más confiables los que se ubican
entre 10.8 y 8.7 Ma (promedio: 10.1 Ma). Estos fechados se aproximan al fechado de 40K/39Ar de 10
Ma obtenido en escorias del Chasiquense típico en la Provincia de Buenos Aires (Schultz et al., 1998).
El Chasiquense típico, unidad de la escala cronológica sudamericana basada en biozonas de mamíferos,
suprayace al “Entrerriense” en su área tipo en tanto que falta en la sección de Paraná donde sobre
el “Entrerriense” (=Formación Paraná) se ubica sedimentos de edad Huayqueriense (=Formación
Ituzaingó), la cual es inmediatamente más joven que el Chasiquense dentro del Tortoniano (véase
más abajo). Los tiempos chasiquenses en el área de Paraná están representados por el hiato de la
discordancia que separa las Formaciones Paraná e Ituzaingó o por la parte superior de la Formación
Paraná (Cione et al., 2000). Si se diera esta última circunstancia, indicaría que el mar “Entrerriense”
perduró más tiempo en la zona de Paraná que en Patagonia o que en el área tipo del Chasiquense.
Esta hipótesis puede estar apoyada por la distinta composición de cetáceos de la Formación Puerto
Madryn y Paraná.
Los seláceos identificados son típicos de los tiempos Mioceno medio a Plioceno (son netamente distintos de aquéllos del “Patagoniense”) pero los cetáceos sugieren mas precisamente una edad Mioceno tardía para la parte superior de la Formación Paraná. El registro de cetáceos Balaenidae y Physeteridae similares a taxones modernos, de Balaenopteridae (desconocidos en capas más viejas que el Mioceno tardío) y la ausencia de taxones comunes con la Formación Gaiman del Mioceno temprano (Cione, en preparación) y la Formación Puerto Madryn del Mioceno Medio-Tardío (Cozzuol, 1993, 1996), sugiere que el extremo superior de la Formación Paraná (al menos en los afloramientos del lecho del Arroyo Ensenada y Villa Urquiza) podría ser de edad tortoniana.
Edad de la Formación Ituzaingó en los alrededores de Paraná
La edad exacta de la Formación Ituzaingó ha sido también largamente debatida. Una discusión detallada de los argumentos basados en las relaciones estratigráficas y en el contenido de la mamalifauna fue abordada por Cione et al. (2000).
Los mamíferos de la Formación Ituzaingó son extremadamente diversos. A primera vista, la coexistencia de tantos taxones diversos podría ser sugestiva de una mezcla secundaria. Algunos autores propusieron que el material proveniente del “Conglomerado osífero” podría haberse redepositado de capas de diferentes edades: Santacrucense, Chasiquense, Huayqueriense, Montehermosense (Bianchini y Bianchini, 1971; Scillato-Yané 1977a, 1980, 1981; Marshall et al., 1983). Por otra parte, muchas de las determinaciones son antiguas y deberían ser revisadas (véase Cione et al., 2000). El trabajo de campo que estamos realizando permitirá clarificar muchos de los problemas al respecto.
Cione et al. (2000), al evaluar la evidencia, destacaron que:
• La edad del “Conglomerado osífero” no parece ser más antigua que Tortoniano por la
relación de suprayacencia (con una discordancia importante) de éste sobre la Formación Paraná.
• El “Conglomerado osífero” posee mayor afinidad con las faunas Chasiquense y Huayqueriense
[Chasiquense, 11 (17); Huayqueriense, 28(32?)]. Descartamos la correlación con Santacrucense,
Mayoense, Laventense y Montehermosense.
• Luego de discutir las afinidades faunísticas, Cione et al. (2000) consideran que el “Conglomerado
osífero” es de edad Huayqueriense.
• La base del Huayqueriense se ubica en 9 Ma (Flynn y Swisher, 1995). De acuerdo a Flynn y
Swisher (1995), las dataciones radioisotópicas disponibles y la ubicación del límite dentro de la
parte temprana del Chron C3Ar, y usando la terminología de Cande y Kent (1995) y Berggren
et al. (1997), la mejor estimación para el límite Huayqueriense/Montehermosense es cerca de
6.8 Ma, y ciertamente más vieja que 6.5 Ma. En realidad, esta estimación no corresponde a las
capas más jóvenes conocidas. Las capas que suprayacen a la Formación Andalhualá posiblemente
son Chapadmalenses (véase Cione y Tonni, 1996). Flynn y Swisher (1995) indican que
el intervalo Huayqueriense/Montehermosense está magnetoestratigráficamente relativamente
bien muestreado en Bolivia y el noroeste de Argentina. Sin embargo, las secciones “Montehermosenses” son Chapadmalenses (Cione y Tonni, 1996). El Huayqueriense es
entonces correlacionado con el Tortoniano de la Escala del Tiempo Geológico (11.0 to 7.3 Ma;
Remane et al., 2000).
Consideramos que el acuífero llamando “Puelchense” (Sala y Auge, 1970; Pascual y Odreman Rivas, 1971) del subsuelo de la región pampeana es correlacionable con la Formación Ituzaingó (ver discusión en Cione y Tonni, 1995; Cione et al., 2000). Esta unidad también suprayace a la Formación Paraná. En el subsuelo del área bonaerense, se han identificado en el “Puelchense” taxones típicos registrados en el “Conglomerado osífero” (Carcharias acutissima, Isurus hastalis, Carcharhinus spp., Myliobatoidei and Sciaenidae; Rusconi, 1948; observacion personal ALC) con vertebrados terrestres (toxodontid y rodents; ver Rusconi, 1948, 1949; Cione y Tonni, 1995; Bond, 1999; Bond et al., 1995; Bond y López, 1998).
La ictiofauna de los alrededores de Paraná
PECES MARINOS
La fauna de elasmobranquios conocida de la Formación Paraná está compuesta por el
odontaspidido Carcharias acutissima (abundante), los carcarrínidos Carcharhinus spp. (abundante) y Physogaleus aduncus, el heterodóntido Heterodontus, el escuatínido Squatina, el lámnido Isurus hastalis,
el otodóntido Carcharocles megalodon, el hemigaleido Hemipristis serra, el escuálido Squalus y por
batoideos Dasyatidae y Myliobatoidei indeterminados (D´Alessandri, 1896; Woodward, 1900; Priem,
1911; Frenguelli, 1920, 1922; Cione, 1978, 1988; Arratia y Cione, 1996; Cione et al., 2000; Figuras 2,3).
La ictiofauna está siendo estudiada, en especial los odontaspididos (Lucifora et al., 2003; Cione et al.,
enviado; Cione et al., en preparación). Algunas de las especies aparecen también en el “Conglomerado
osífero”, seguramente retrabajadas de los sedimentos marinos.
Varios peces óseos fueron reportados. El espárido Chrysophrys sp., Sparidae indet., y el lábrido Protautoga longidens Alessandri, 1896 fueron descriptos y figurados (Alessandri, 1896). El material fue perdido durante la Primera Guerra Mundial en Turín, Italia. Sin embargo a partir de las ilustraciones se puede observar que el material no pertenece ni a Sparidae ni a Labridae. El material identificado como Sparidae indet. (Alessandri, 1896) es muy similar a placas faríngeas inferiores del esquiénido Pogonias cromis. Nosotros hemos encontrado restos de esquiénidos tanto en la Formación Paraná como en el “Conglomerado osífero” (Cione y Torno, 1984; Cione et al., 2000; Figura 4).
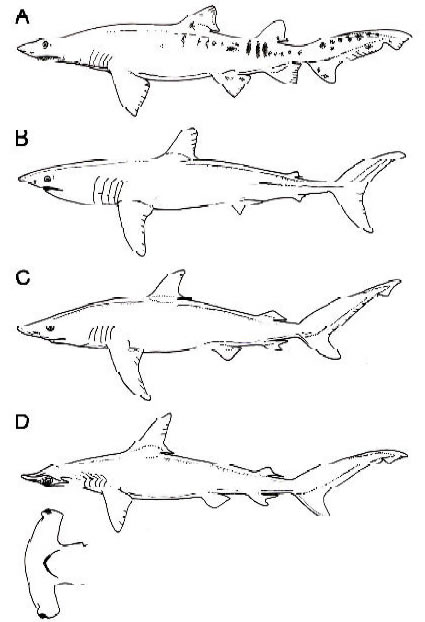
Fig. 3. Representantes vivientes de géneros de neoseláceos que se registran en la Formación Paraná. A Carcharias taurus. B, Isurus oxyrinchus. C. Carcharhinus obscurus. D. Sphyrna zygaena. Todos modificados de Bigelow y Schroeder (1948). No están a escala.
PECES CONTINENTALES
Rayas del género Potamotrygon de la familia de agua dulce Potamotrygonidae fueron mencionados
por Ameghino (1898) aunque no los describió ni figuró (Figura 3). Más recientemente, Deynat y
Brito (1994) asignaron grandes escudos dérmicos descriptos por Larrazet (1886) con procedencia de
Paraná a la familia Potamotrygonidae. Escudos dérmicos similares fueron registrados por nosotros
tanto en la marina Formación Paraná como en el continental “Conglomerado osífero.” Sin embargo,
parte del material probablemente corresponda a otras familias de rayas.
Numerosos especímenes de siluriformes de agua dulce y caraciformes se registran en la Formación Ituzaingó. En la Formación Paraná han aparecido bagres marinos (Familia Ariidae). De hecho, estos peces han sido parcialmente descriptos y figurados por varios autores desde el siglo XIX (Bravard, 1858; Ameghino, 1898; Woodward, 1900; Priem, 1911; Figura 4). Sin embargo, ellos no han sido aun adecuadamente estudiados (véase comentarios en Cione, 1978, 1986; Arratia y Cione, 1996).
El bagre Silurus agassizi Bravard, 1858 no fue descripto ni ilustrado y debe ser considerado nomen nudum (Cione, 1986). La asignación de un cráneo de bagre a S. agassizi por Frenguelli (1920) es consecuentemente inválida. Hemos observado que siluriformes de diferentes familias están presentes en el material del “Conglomerado osífero”. Algunos caraciformes como el serrasálmido Colossoma macropomum se encuentran en la Formación Ituzaingó (Cione, 1986). Esta especie ha sido también identificada en el Grupo La Venta (Mioceno Medio de Colombia; Lundberg et al., 1986, 1988). Los registros de la Formación Ituzaingó Formation y el Grupo La Venta corroboran las relaciones biogeográficas postuladas para otros elementos faunísticos (por ejemplo, cocodrilos, triquéquidos, cetáceos). Un nuevo género de Characidae representa un elemento intermedio entre los representantes más generalizados y las pirañas (Cione et al., 2003).
Los “holósteos” Gynglimodi fueron prácticamente ecuménicos en el Cretácico. En el Cenozoico
redujeron notablemente su distribución y no habitaron más Sudamérica. Sin embargo, D´Alessandri
(1896) asignó un diente al Ginglymodi Lepisosteus. El material en realidad corresponde a un Crocodylia
indeterminado (Cione, 1986).
Dientes caniniformes de la familia Cynodontidae (Characiformes), similares a los de los géneros Hydrolicus, Cynodon y Raphiodon, fueron descriptos del “Conglomerado Osífero” (Cione y Casciotta, 1997). Sin embargo el tamaño y posición de la sierra sobre los bordes cortantes es diferente de la registrada en especies de estos géneros. Un gran diente, probablemente corresponda a un especimen de más de 1000 mm de longitud estándar.
Existen numerosos restos de Siluriformes, Characiformes y Perciformes que están en proceso de estudio. Es posible sostener que la diversidad existente en agua dulce en el área ha sido sólo vislumbrada.
Biogeografía
PECES MARINOS
Los géneros de peces marinos conocidos de la Formación Paraná y que tienen representantes
vivientes habitan la actual Provincia biogeográfica Argentina (sensu López, 1964; ver Cione, 1978,
1988; Figura 2) y algunos aguas más cálidas (Hemipristis). Los peces integran ambientes neríticos,
probablemente de plataforma interna. No se hallaron aún peces típicamente oceánicos (por ejemplo,
istiofóridos, los denominados peces vela).
Es notable la presencia de tiburones heterodóntidos, los cuales no habitan en la actualidad el
Océano Atlántico (Cione y Azpelicueta, 2002). En el Oligoceno superior-Mioceno medio de Patagonia
se han registrado otros peces que no habitan el Atlántico o al menos el Atlántico sur tales como los
tiburones pristiofóridos y los teleósteos oplegnátidos (Cione y Expósito, 1980; Cione y Azpelicueta,
2002). En el Terciario, especies de Heterodontus y pristiofóridos tuvieron una distribución casi ecuménica (Cappetta, 1987; Cione, 1988). Hoy las especies de Heterodontus viven en el Pacífico (Compagno,
2001; Compagno et al., 2005), mientras que los pristiofóridos no habitan en la actualidad las costas
de los océanos Pacífico y Atlantico en Sudamérica (Compagno, 2001). En el presente, una pequeña
población de la especie endémica Pristiophorus schroederi habita en el noroeste del Atlántico (Springer
y Bullis, 1960). Ambos grupos redujeron su distribución después del Mioceno. El hallazgo de
Pristiophorus en la Formación Puerta del Diablo de Patagonia y Heterodontus en la Formación Paraná
constituye el último registro de este género en el Atlántico sur y en Atlántico respectivamente.
Ambos representan un ejemplo de extirpación en un área particular y persistencia en otra (Cione y
Azpelicueta, 2002).
Los cetáceos Pontoporiidae fueron también comunes en el norte del Pacífico Sur (norte de Chile y sur de Perú, Cozzuol, 1996), en el Pacífico Norte (Barnes, 1985) y Atlántico Norte. La presencia de pontopóridos en la Formación Paraná sugieren una biogeografica conexión con aquellas regiones, en tiempos en los cuales el puente de America Central estaba aun abierto.
Dioplotherium, aparentemente en aquellos depósitos, fue también hallado en el norte de Brazil (Formación Pirabas, de Toledo y Domning, 1991) y en el Pacífico norte y Atlántico norte (Domning, 1989). Se piensa que los balenoptéridos se originaron en el Atlantico tropical no antes del Mioceno medio tardío, pero ellos se encuentran distribuidos alrededor del mundo en el Mioceno tardío como es el caso de los registros de Formación Paraná.
Si las afinidades de Monachus con Properiptychus argentinus son confirmadas, este grupo posee una distribución tropical.
Consecuentemente, los mamíferos acuáticos indican una fuerte conexión con localidades más norteñas. Para explicar esto no es necesario apelar a conexiones a través del continente ya que es suficiente la conexión de oceánica a lo largo de la costa continental.
PECES CONTINENTALES
La Región Neotropical se encuentra dividida en dos subunidades fenéticas de acuerdo a la
distribución actual de los peces: las subregiones Brasílica y Austral (Ringuelet, 1975; Arratia et al.,
1983). La subregión Austral incluye Patagonia, Cuyo, y Sur y centro de Chile (Ringuelet, 1975; Arratia
et al., 1983; Arratia, 1997). El límite norte de la subregión Austral se encuentra definido en Argentina
por las cuencas de los ríos San Juan-Desaguadero-Curacó-Colorado. La Subregión Brasílica ocupa el
resto de Sudamérica y América Central. La Subregion Brasílica es el área más rica en diversidad de
peces y la Subregión Austral una de las menos diversas en el mundo (menos de 25 species; Ringuelet,
1975; Almirón et al., 1997; Arratia, 1997). La diversidad brasílica disminuye notablemente al sur del
Río de la Plata, aún dentro de la subregión.
Los peces de agua dulce del Mioceno de Paraná son típicamente brasílicos y no fue hallado ningún taxón austral (Figura 4).
Los peces, cocodrilos, cetáceos, y sirénidos del “Conglomerado osífero” se encuentran relacionados con aquéllos del Mioceno tardío de las capas del Mioceno de La Venta, en Colombia (Langston, 1965; Langston y Gasparini, 1997; Cione, Lundberg y Machado-Allison, en preparación), las capas del Mioceno tardío de Perú y del Acre brasileño (Rancy et al., 1989; Bocquentin y Souza Filho, 1990; Frailey, 1986; Gasparini, 1996) y las capas del Mioceno tardío-Plioceno temprano de Urumaco, Venezuela (Buffetaut, 1982; Gasparini, 1996). La Venta, Acre y Paraná comparten Caiman, Mourasuchus y Gryposuchus. Acre y Paraná también comparten Ischyrorhynchus, Saurocetes y Ribodon.
En la actualidad, la cuenca del Paraná-Uruguay-Plata está prácticamente aislada de la cuenca del Amazonas. La presencia de vertebrados acuáticos de agua dulce similares con las capas del Mioceno de Colombia (actual cuenca del Magdalena), Venezuela y el Amazonas brasileño indica relaciones hidrográficas que no existen hoy. La presencia de cetáceos ínidos, sirenios triquéquidos y una alta diversidad de cocodrilos también sugieren un clima más cálido que el que existe en el presente.
Ambiente
MARINO
El nivel alto del mar en el Mioceno medio y quizás tardío permitió la ambientes de cierta
salinidad en la planicie chacopampeana al menos hasta Paraguay y Bolivia (Uliana y Biddle, 1988;
Marshall et al., 1993; véase Aceñolaza, 2004). En el sur, la influencia marina estuvo restringida al
noreste de la Patagonia. Una ictiofauna miocena templada se registra en el extremo superior de la
Formación Paraná. El conjunto ictiofaunístico mioceno de Paraná es diferente al patagónico, pero es
similar al presente a latitudes similares o más norteñas en la costa atlantica del sur del Brazil (Cione,
1978; Cione et al., 2000). Mientras que carcarrínidos, hemigaleidos, y odontaspididos dominan en
Paraná, la ictiofauna patagónica esta dominada por lámnidos (Cione, 1978, 1988; Cione y Tonni,
1981; Perea et al., 1985; Perea y Ubilla, 1989, 1990). Los cetáceos de la Formación Paraná también
indican temperaturas no muy diferentes de las que se presentan a la misma latitud o más cálidas. Los
sirenios, los pinnípedos y los invertebrados sugieren aguas más cálidas para el área durante el
Mioceno, las que podrían haberse extendido hasta el norte de la Patagonia teniendo en cuenta los
invertebrados (Del Río, 1988; 2000; Cione et al., 2000).
La asociación de elasmobranquios presentes en la Formación Paraná sugiere una salinidad marina normal en los niveles y el área en que está presente (desde la Bajada de la Celina, cerca de Villa Urquiza hasta Victoria y en el subsuelo del noreste de la provincia de Buenos Aires).
CONTINENTAL
Todos los restos de vertebrados terrestres de la Formación Ituzaingó se encuentran desarticulados.
Sin embargo esto es usual en depósitos fluviales. Los únicos restos de vertebrados articulados son
algunos peces de agua dulce muy bien preservados en nódulos. Notablemente, muchos huesos
están escasamente erosionados. Nosotros consideramos poco probable que el material provenga de
otras unidades terrestres (las cuales son desconocidas en el área; ver también Cozzuol, 1993). Más
aún, la cuenca fluvial se desarrolló sobre capas marinas y no hay duda que alguno (no necesariamente
todos) de los restos marinos presentes en el “Conglomerado osífero” fueron retrabajados de la
Formación Paraná. Recientemente han aparecido algunos restos de mamíferos terrestres en la
Formación Paraná en Aldea Brasilera (inédito).
No se registraron dipnoos en el área. Tomando en cuenta la buena preservación de los dientes de dipnoos, la ausencia de estos peces podría estar relacionada a la ausencia de biestacionalidad o de ambientes adecuados.
La ubicación paleogeográfica de Paraná en el cinturón subtropical a templado es también confirmada por la ausencia de crocodilidos, podocnémidos, y primates. Actualmente, los cocodrilos sudamericanos habitan exclusivamente regiones tropicales. Sin embargo la ausencia de primates podría deberse a la preservación insuficiente.
La diversidad taxonómica de los reptiles, especialmente los crocodilos del “Conglomerado osífero” sugieren una variedad de paleoambientes. La predominancia de aves acuáticas en el “Conglomerado osífero” señalan la presencia de áreas bajas boscosas y pantanosa lo largo de los bancos del río. Este tipo de ambientes es tambien esencial para los triquéquidos y los ínidos, los cuales necesitan lagos asociados a ríos principales para vivir, comer y reproducirse.
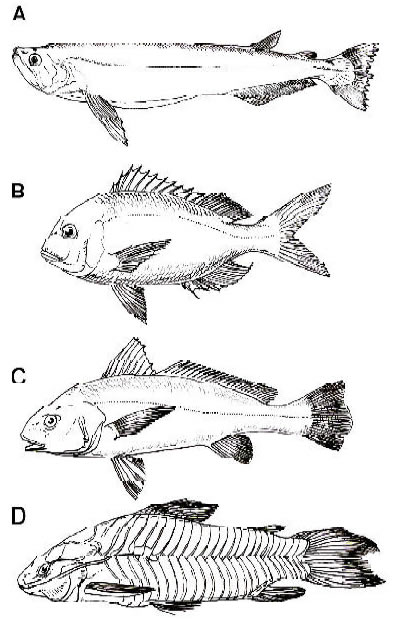
Fig. 4. Representantes vivientes de géneros de teleósteos que se registran en la Formación Ituzaingó. A, Rhaphiodon vulpinus (modificado de Ringuelet et al., 1967). B, Pagrus pagrus (modificado de Cousseau y Perrotta, 2000). C, Micropogonias furnieri (modificado de Cousseau y Perrotta, 2000). D. Hoplosternum littorale (modificado de Casciotta et al., 2003). No están a escala.
Agradecimientos
Los autores agradecen a las siguientes instituciones y personas:
Al Consejo Nacional de Investigaciones Científicas y Tecnológicas, a la Agencia Nacional de Promoción Científica y Tecnológica, a la Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, y a la Universidad Nacional de La Plata, por su apoyo financiero permanente.
Al Centro de Investigaciones Científicas y Transferencia de Diamante, por asistencia en el área de Paraná.
A Florencio Aceñolaza por su invitación a participar en el presente volumen. A C. Ceruti, por ayuda en el Museo de Paraná. A José Laza, por sus excelentes preparaciones del material colectado. A Carlos Tremouilles y Jorge González, por dibujos. A Carlos Steger, por prestarnos algunos materiales de su colección.
Bibliografía
Aceñolaza, F.G. 1976. Consideraciones bioestratigráficas sobre el Terciario marino de Paraná y alrededores. Acta Geológica Lilloana 13: 91-107.
Aceñolaza, F.G. 2004. Paleobiogeografía de la Región Mesopotámica. In: Aceñolaza, F.G. (Coord.) Temas de la Biodiversidad del Litoral fluvial argentino. Miscelánea Insugeo 12:25-30.
Aceñolaza, F.G. y Aceñolaza, G. 1999. Trazas fósiles del Terciario marino de Entre Ríos (Formación Paraná, Mioceno medio), República Argentina. Boletín de la Academia Nacional de Ciencias, Córdoba 64: 209-233.
Aceñolaza, F.G. y J.M. Sayago. 1980. Análisis preliminar sobre la estratigrafía, morfodinámica y morfogénesis de la región de Villa Urquiza, provincia de Entre Ríos. Acta Geológica Lilloana 15: 139-154.
Alessandri, G. d´. 1896. Ricerche sui pesci fossili de Paraná. Atti della Reale Academia di Scienze di Torino 31: 1-17.
Almirón, A., Azpelicueta, M.M., Casciotta, J. y López Cazorla, A. 1997. Ichthyogeographic boundary between the Brazilian and Austral subregions in South America, Argentina. Biogeographica 73: 23-30.
Ameghino, F. 1883a. Sobre una colección de mamíferos fósiles del Piso Mesopotámico de la Formación Patagónica recogidos en las barrancas del Paraná por el Profesor Pedro Scalabrini. Boletín de la Academia Nacional de Ciencias de Córdoba (República Argentina) 5 (entrega 1a): 101-116.
Ameghino, F. 1883b. Sobre una nueva colección de mamíferos fósiles recogidos por el Profesor Scalabrini en las barrancas del Paraná. Boletín de la Academia Nacional de Ciencias de Córdoba 5: 112-113.
Ameghino, F. 1885. Nuevos restos de mamíferos fósiles oligocenos recogidos por el profesor Pedro Scalabrini y pertenecientes al Museo Provincial de la ciudad de Paraná. Boletín de la Academia Nacional de Ciencias de Córdoba 8: 3-207.
Ameghino, F. 1886. Contribución al conocimiento de los mamíferos fósiles de los terrenos Terciarios antiguos del Paraná. Boletín de la Academia Nacional de Ciencias de Córdoba (República Argentina) 9: 3-226.
Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias de Córdoba. 6: XXXII + 1028, Atlas: 98 láms.
Ameghino, F. 1898. Sinopsis geológico-paleontológica. In: 2do Censo de la República Argentina. I. Territorio, Buenos Aires: 1-500.
Ameghino, F. 1906. Les Formations Sedimentaires du Crétacé Supérieur et du Tertiaire de Patagonie avec un paralléle entre leurs faunes mammalogiques et celles de l’ancien continent. Anales del Museo Nacional de Buenos Aires 15 (ser. 3a, 8): 1-568.
Arratia, G., Peñafort, B. y Menu-Marque, S, 1983. Peces de la Región Sureste de los Andes y sus probables relaciones biogeográficas actuales. Deserta 7: 48-107.
Arratia, G. y Cione, A.L. 1996. The fossil fish record of Southern South America. In: G. Arratia (ed.). Contributions of Southern South America to Vertebrate Paleontology, München Geowissenschaft Abhandlungen, 30, pp. 9-72.
Arratia, G. 1997. Brazilian and Austral freshwater fish faunas of South America. A contrast. In: Tropical biodiversity and systematics. Proceedings of the International Symposium on Biodiversity and Systematics in Tropical Ecosystems, Bonn, 1994: 179-186.
Berggren, W.A., Hilgen, F.J., Langereis, C.G., Kent, D., Obradovich, J., Raffi, I., Raymo, M.E., y Shackleton, N.J. 1997. Late Neogene chronology: new perspectives in high-resolution stratigraphy. Bulletin of the Geological Society of America 107: 1272-1287.
Bigelow, H.B. y Schroeder, W.C. 1948. Fishes of the Western North Atlantic. Memoirs of the Sears Foundation for Marine Research 1: 59-552.
Bianchini, L.H. Delupi de, y Bianchini, J.J. 1971. Revisión de los Proterotheriinae (Mammalia, Litopterna) del Mesopotamiense. Ameghiniana 8: 1-24.
Bocquentin, J. y Souza Filho, J. 1990. O crocodiliano sud-americano Carandaisuchus como sinonimia de Mourasuchus (Nettosuchidae). Revista Brasileira de Geociencias 20: 230-233.
Bond, M. 1999. Quaternary native ungulates of Southern South America. A synthesis. In: E. P. Tonni y A. L. Cione (eds.). Quaternary vertebrate paleontology in South America. Quaternary of South America 12. Special Volume, pp. 177-205.
Bond, M. y López, G.M. 1998. Notopachyrukhos tambuttoi Rusconi, 1933, un Interatheriidae (Mammalia, Notoungulata) del Puelchense de la provincia de Buenos Aires. Actas del X Congreso Latinoamericano de Geología y VI Congreso Nacional de Geología Económica 1: 280-283. Buenos Aires.
Bond, M.; Cerdeño, M.E. y López, G.M. 1995. Ungulados Nativos Sudamericanos. In: M. T Alberdi, G. Leone y E. P. Tonni (eds.). Evolución Biológica y Climática de la región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo Occidental. Monografías del Museo Nacional de Ciencias Naturales, Madrid, CSIC, pp. 257-275.
Bondesio, P., Laza, J., Scillato Yané, G., Tonni, E.P. y Vucetich, M.G. 1980. Estado actual del conocimiento de los vertebrados de la Formación Arroyo Chasicó (Plioceno temprano) de la provincia de Buenos Aires. Actas II Congreso Argentino de Paloentología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología 3: 101-127.
Bravard, A. 1858. Monografía de los terrenos terciarios del Paraná. (Reimpresión facsimilar con prólogo de F.G. Aceñolaza, Imprenta del Congreso de la Nación, Buenos Aires, 1995,V-XII + 1-107 pp.).
Buffetaut, E. 1982. Systématique, origine et évolution des Gavialidae Sud-Américains. Géobios 6: 127-140.
Cande, S.C. y Kent, D. 1995. Revised calibration of the Geomagnetic Polarity Time Scale for the Late Cretaceous and Cenozoic. Journal of Geophysical Research 100: 6093-6096.
Cappetta, H. 1987. Chondrichthyes II. In: H.-P. Schultze (ed.). Handbook of Paleoichthyology. Gustav Fischer Verlag, Stuttgart, New York, pp. 1-193.
Casciotta, J.R., Almirón, A. y Bechara, J. 2003. Los peces de la laguna Iberá. Ediciones Al Margen, La Plata, 203 pp.
Castellanos, A. 1936. Los sedimentos del Pampeano inferior y del Araucanense en el valle de los Reartes (Sierra de Córdoba). Publicaciones de la Facultad de Ciencias Matemáticas, Físicoquímicas y Naturales 6: 1-16.
Chebli, G.A., Tófalo, O. y Turzzini, G. 1989. Mesopotamia. In: G.A. Chebli y L. Spalletti (eds.). Cuencas sedimentarias argentinas. San Miguel de Tucumán, Universidad Nacional de Tucumán, Instituto de Correlación Geológica, Serie Correlación Geológica 6: 79-100.
Cione, A.L. 1978 Aportes paleoictiológicos al conocimiento de la evolución de las paleotemperaturas en el área austral del América del Sur durante el Cenozoico. Aspectos zoogeográficos y ecológicos conexos. Ameghiniana15: 183-208.
Cione, A.L. 1986. Los peces continentales del Cenozoico de Argentina. Su significación paleoambiental y paleobiogeográfica. Actas del III Congreso Argentino de Paleontología y Bioestratigrafía 2: 101-106.
Cione, A.L. 1988. Los peces de las formaciones marinas del Cenozoico de Argentina. Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, pp. 588.
Cione, A.L. y M.M. Azpelicueta. 2002. An oplegnathid fish from the Miocene of Patagonia. Geobios 35:367-373.
Cione, Alberto Luis, Azpelicueta, M.M., Bond, M., Carlini, A., Casciotta, J.R., Cozzuol, M.A., Fuente, M. de la,
Gasparini, Z., Goin, F., Noriega, J., Scillato-Yané, G.J., Soibelzon, L., Tonni, E.P., Verzi, D., Vucetich, M.G. 2000. The Miocene vertebrates from Paraná, eastern Argentina. In: Aceñolaza, F.G. & R. Herbst (eds.). El Neógeno de
Argentina. Serie Correlación Geológica 14: 191-237. http://www.unt.edu.ar/fcsnat/INSUGEO/Libro_neogeno/
13_miocene_vertebrates_from_entre_r.htm
Cione, A.L, M.M. Azpelicueta, J. R. Casciotta y M.T. Dozo. 2005. Tropical freshwater teleosts form Miocene beds of eastern Patagonia, southern Argentina. Geobios 38: 29-42.
Cione, A.L. y Expósito, S. 1980. Chondrichthyes del “Patagoniano” s.l. de Astra, Golfo de San Jorge, provincia de Chubut, Argentina. Su significado paleoclimático y paleobiogeográfico. Actas del II Congreso Argentino de Paleontología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología 2: 275-290. Buenos Aires.
Cione, A.L. y Torno, A.. 1984. Descripción y comparación de peculiares estructuras en la aleta dorsal de Pogonias cromis (Perciformes, Sciaenidae) y de una especie de la misma familia del Terciario tardío de Entre Ríos, Argentina. Resúmenes VI Jornadas Argentinas de Zoología: 178.
Cione, A.L. y Casciotta, J.R. 1995. Freshwater teleostean fishes from the Miocene of the Quebrada de la Yesera, Salta, Northwestern Argentina. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 196: 377-394.
Cione, A.L. y Tonni, E.P. 1995. Chronostratigraphy and “Land-mammal ages” in the Cenozoic of southern South America: principles, practices, and the “Uquian” problem. Journal of Paleontology 69: 135-159.
Cione, A.L. y Tonni, E.P.1996. Reassesment of the Pliocene-Pleistocene continental time scale of southern South America. Correlation of the Chapadmalalan with Bolivian sections. Journal of South American Earth Sciences 9: 221-236.
Cione, A.L. y Casciotta, J.R. 1997. Miocene cynodontids (Osteichthyes: Characiformes) from Paraná, central eastern Argentina. Journal of Vertebrate Paleontology 17: 616-619.
Cione, A.L., Machado-Allison, A., Lundberg, J. y Dahdul, W. 2003. A new genus and species of Serrasalminae (Characidae, Teleostei) from the upper Miocene of Argentina. Abstracts of the Joint Meeting of Ichthyologists and Herpetologists, Manaus, Brazil, June 26, 2003.
Cione, A.L., J. Mennucci, F. Santalucita y C.Acosta Hospitaleche. Local extinction of genus Carcharias (Elasmobranchii, Odontaspididae) in the eastern Pacific Ocean.. Revista Geológica de Chile.
Compagno, L. 2001. Sharks of the World: An annotated and illustrated catalogue of shark species known to date. Vol. 2. Bullhead, mackerel and carpet sharks (Heterodontiformes, Lamniformes and Orectolobiformes). Fao Species Catalogue for Fishery Purposes 1(2). 325 pp., Rome.
Compagno, L., Dando, M. y Fowler, S. 2005. Sharks of the World. Collins Field Guide, Londres, 369 pp.
Cousseau, M.B. y Perrotta, R.G. 2000. Peces marinos de Argentina. Inidep, Mar del Plata, 167 pp.
Cozzuol, M.A. 1985. The odontoceti of the «Mesopotamiense» of the Paraná River Ravines. Systematic Review. In: G. Pilleri (ed.). Investigations on Cetacea 17, pp. 39-54.
Cozzuol, M.A. 1989. Una nueva especie de Saurodelphis Burmeister, 1891 (Cetacea: Iniidae) del “Mesopotamiense” (Mioceno tardío-Plioceno temprano) de la provincia de Entre Ríos, Argentina. Ameghiniana 25: 39-45.
Cozzuol, M.A. 1993. Mamíferos acuáticos del Mioceno medio y tardío de Argentina. Sistemática, evolución y biogeografía. Tesis doctoral, Universidad Nacional de La Plata, Facultad de Ciencias Naturales y Museo de La Plata, inédita.
Cozzuol, M.A. 1996. The record of the aquatic mammals in southern South America. In: G. Arratia, (ed.). Contributions to southern South America to Vertebrate Paleontology, Münchner Geowissenschaftliche Abhandlungen, Reihe A, Geologie und Paläontologie 30: 321-342.
Darwin, C. 1839. Voyage of the Beagle. Journal of researches into the Geology and Natural History of the various Countries visited by H.M.S Beagle. Prometheus Books, London, 547 p.
Deynat, P. P. y Brito, P. M. 1994. Révision des tubercules cutanés de raies (Chondrichthyes, Batoidea) du bassin du Paraná, Tertiaire d’Amerique du Sud. Annales de Paléontologie 80: 237-251.
Doering, A. 1882. Informe oficial de la Comisión científica agregada al Estado Mayor General de la expedición al Río Negro. Geología. Buenos Aires: 299-530.
Domning, D.P. 1982. Evolution of manatees: a speculative history. Journal of Paleontology 56:599-619.
Feruglio, E. 1949. Descripción geológica de la Patagonia 2. Yacimientos Petrolíferos Fiscales, Buenos Aires. 349 p.
Flynn, J.J. y Swisher, C.C. 1995. Cenozoic South American Land-mammal ages: correlation to global geochronologies. In: W.A. Berggren, D.V. Kent, y J. Handerbol (eds.). Geochronology, Time scales, and Correlation: Framework for a Historical Geology. SEPM Special Publication, 54: 317-333.
Frailey, C.D. 1986. Late Miocene and Holocene mammals, exclusive of the Notoungulata, of the rio Acre region, western Amazonia. Contribution on Science 374: 1-46.
Frenguelli, J. 1920. Contribución al conocimiento de la geología de Entre Ríos. Boletín de la Academia Nacional de Ciencias de Córdoba (República Argentina) 24: 55-256.
Frenguelli, G. 1922. Sulla contemporanea presenza del Carcharodon megalodon Ag. e del Carcharodon rondeleti M.et H. nel Paranense superiore de Entre Ríos (Rep. Argentina). Bolletino de la Societá Geologica Italiana 41: 50-52.
Frenguelli, G. 1930. Las guayquerías de San Carlos en la provincia de Mendoza. Publicaciones del Departamento de Extensión Universitaria, Universidad Nacional del Litoral 9: 1-54.
Galván, A.F. y O.J. Ruiz Huidobro. 1965. Geología del valle de Santa María. Estratigrafía de las formaciones mesozoico-terciarias. Actas de las Segundas Jornadas Geológicas Argentinas 3:217-230. Córdoba.
Gasparini, Z. 1996. Biogeographic Evolution of the South American Crocodilians. In: G. Arratia (ed.). Contributions of Southern South America to Vertebrate Paleontology. Müchner Geowissenschaftliche Abhandlungen 30, pp. 159-184.
Guler, M.V., Guerstein, G. y Malumián, N. 2002 - Bioestratigrafía de la Formación Barranca Final, Neógeno de la Cuenca del Colorado, Argentina. Ameghiniana, 39: 103-110.
Handerbol, J., Thierry, T., Farley, M.B., Jacquin, R., Graciansky, P. y Vail, P. 1998. Mesozoic and Cenozoic sequence chronostratigraphic framework of European basins. In: Graciansky, P.-C., J. Handerbol, Jacquin, T. y Vail, P.R. (eds.). Mesozoic and Cenozoic sequence stratigraphy of European basins. Society for Sedimentary Geology Special Publication 60:3-15.
Iriondo, M.H. 1980. El cuaternario de Entre Ríos. Revista de la Asociación de Ciencias Naturales del Litoral 11: 125-141.
Iriondo, M.H. y E.D. Rodríguez. 1973. Algunas características sedimentológicas de la Formación Ituzaingó entre La Paz y Pueblo Brugo (Entre Ríos). Actas del V Congreso Geológico Argentino 1: 317-331.
Kraglievich, J.L. 1952. El perfil geológico de Chapadmalal y Miramar, Provincia de Buenos Aires. Revista Museo Municipal Ciencias Naturales y Tradicional Mar del Plata 1:8-37.
Langston, W. 1965. Fossil crocodilians from Colombia and the Cenozoic history of the Crocodilids in South America. University of California Publications in Geological Sciences 52: 1-157.
Langston, W. y Gasparini, Z. 1997. Crocodilians, Gryposuchus, and the South American Gavials. In: R. Kay, R. Madden, R. Cifelli y J. Flynn (eds.). Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta. Smithsonian Institution Press 8: 113-154.
Larrazet, A. 1886. Des pièces de la peau de quelques Sélaciens fossiles. Bulletin de la Société Géologique de France 14:259-261.
López, R. 1964. Problemas de la distribución geográfica de los peces marinos suramericanos. Boletín del Instituto de Biología Marina 7: 57-63.
Lucifora, L.O., A.L. Cione, R.C. Menni y A.H. Escalante 2003. Tooth row counts, vicariance, and the distribution of the sand tiger shark, Carcharias taurus. Ecography 26: 567-572.
Lundberg, J. 1997. Fishes of the La Venta fauna: additional taxa, biotic and paleoenvironmental implications. In: R.F. Kay, R. H.Madden, R. L. Cifelli, y J. J. Flynn (eds.). Vertebrate paleontology in the Neotropics: The Miocene fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, D.C, pp. 67-91.
Lundberg, J., Machado-Allison, A. y Kay, R. 1986. Miocene characid fishes from Colombia: evolutionary stasis and extirpation. Science 234: 208-209.
Lundberg, J.G., Linares, O.J., Antonio, M.E. y Nass, P. 1988. Phractocephalus hemiliopterus (Pimelodidae, Siluriformes) from the upper Miocene Urumaco Formation, Venezuela: a further case of evolutionary stasis and local extinction among South American fishes. Journal of Vertebrate Paleontology 8: 131-138.
Marengo, H. 2000. Rasgos micropaleontológicos de los depósitos de la transgresión Entrerriense-Paranense en la cuenca Chaco-Paranense y noroeste argentino. Serie de Correlación Geológica, 14: 29-45.
Marshall, L.G. 1985. Geochronology and Land-mammal biochronology of the transamerican faunal interchange. In: The Great American Interchange. D. Stehli y S.D. Webb. Plenum Press, p. 49-78.
Marshall, L.G. y Patterson, B. 1981. Geology geochronology of the mammal-bearing Tertiary of the valle de Santa María and río Corral Quemado, Catamarca province, Argentina. Fieldiana Geology 9: 1-80.
Marshall, L., Hoffstetter, R. y Pascual, R.. 1983. Mammals and stratigraphy: geochronology of the continental mammal-bearing Tertiary of South America. Palaeovertebrata Mémoire Extraordinaire: 1-93.
Marshall, L.G., Sempere, T. y Gayet, M. 1993. The Petaca (Late Oligocene-Middle Miocene) and Yecua (Late Miocene) formations of the Subandean Chaco basin, Bolivia, and their tectonic significance. Documents du Laboratoire de Géologie, Lyon 125: 291-301.
Marshall, L., Berta, A., Hoffstetter, R., Pascual, R., Reig, O., Bombin, M. y Mones, A.. 1984. Mammals and stratigraphy: geochronology of the continental mammal- bearing Quaternary of South America. Palaeovertebrata Mémoire Extraordinaire 1-76.
Moussy, M. de. 1857. Cuadro general de la Ciudad del Paraná. Nacional Argentino. 161, 162, 163, 164, Paraná.
Moussy, M. de. 1860. Description Physique, Géographique et Statistique de la Conféderation Argentine. I-III + Atlas.
Noriega, J.I. 1995. The avifauna from the “Mesopotamian” (Ituzaingó Formation: Upper Miocene) of Entre Ríos Province, Argentina. Courier Forschungsinstitut Senckenberg 181: 141-148.
North American Commision on Stratigraphic Nomenclature. 1983. North American Stratigraphic Code. Bulletin American Association of Petroleum Geologists 67: 841-875.
Orbigny, A d’. 1842. Voyage dans l’Amerique méridionale. P. Bertrand, Paris, pp.188.
Pascual, R. y Odreman Rivas, O. 1971. Evolución de las comunidades de los vertebrados del Terciario argentino. Los aspectos paleozoogeográficos y paleoclimáticos relacionados. Ameghiniana 8: 372-412.
Pascual, R., Ortega Hinojosa, E., Gondar, D. y Tonni E.P. 1965. Las edades del Cenozoico mamalífero de la Argentina, con especial atención a aquellas del Territorio Bonaerense. Anales de la Comisión de Investigación Científica 6:165-193.
Priem, F. 1911. Poissons fossiles de la République Argentine. Bulletin de la Société Belge de Géologie, Paléontologie et Hydrologie: 329-340.
Rancy, A., Boquentin Villanueva, J., Pereira de Souza Filho, J., Santos, J.C.R. y Negri, F.R. 1989. Lista preliminar da fauna do Neógeno da regiáo oriental do estado do Acre, Brasil (Material depositado em Rio Branco). VII Jornadas Argentinas de Paleontología de Vertebrados, Buenos Aires, Ameghiniana 26: 249.
Reig, O.A., 1957. Sobre la posición sistemática de “Zygolestes paranensis” Amegh. y de “Zygolestes entrerrianus” Amegh., con una reconsideración de la edad y correlación del “Mesopotamiense”. Holmbergia 5: 209-226.
Reig, O. 1981. Teoría del origen y desarrollo de la fauna de mamíferos de América del Sur. Monographie Naturae 1: 1-1182.
Remane, J., Faure-Muret, A. y G.S. Odin, G.S. 2000. Activo en 2005. International Stratigraphic Chart. ftp:// ftp.iugs.org/pub/iugs/iugs_intstratchart.pdf
Riggs, E.S. y Patterson, B. 1939. Stratigraphy of Late-Miocene and Pliocene deposits of the Province of Catamarca (Argentina) with notes on the faunae. Physis 14: 143-162.
Ringuelet, R. 1975. Zoogeografía y ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur 2: 1-122.
Río, C.J. del. 1988. Bioestratigrafía y cronoestratigrafía de la Formación Puerto Madryn (Mioceno medio) Provincia del Chubut, Argentina. Anales de la Academia Nacional de Ciencias Exactas, Físicas y Naturales 40: 231-254.
Río, C.J. del. 1990 - Composición, origen y significado paleoclimático de la malacofauna “Entrerriense” (Mioceno medio) de la Argentina. Anales de la Academia Nacional de Ciencias Exactas, Físicas y Naturales, 42: 207-226.
Río, C.J. del. 1991. Revisión sistemática de los bivalvos de la Formación Paraná (Mioceno medio), provincia de Entre Ríos, Argentina. Monografías de la Academia Nacional de Ciencias Exactas, Físicas y Naturales 7: 11-90.
Río, C.J. del, Martínez, S. y Scasso, R. 2001 - Nature and origin of spectacular marine Miocene shell beds of northeastern Patagonia (Argentina): paleoecological and bathymetric significance. Palaios, 16: 3-25.
Rusconi, C. 1948. El Puelchense de Buenos Aires y su fauna (Plioceno medio). (Primera parte). Publicaciones del Instituto de Fisiografía y Geología, Universidad Nacional del Litoral 33: 1-99.
Rusconi, C. 1949. El Puelchense de Buenos Aires y su fauna (Plioceno medio). (Segunda parte). Publicaciones del Instituto de Fisiografía y Geología, Universidad Nacional del Litoral 36: 100-242.
Sala, J.M. y Auge, M.P. 1970. Algunas consideraciones geohidrológicas del Noreste de la provincia de Buenos Aires. Actas IV Jornadas Geológicas Argentinas 2: 321-336.
Santa Cruz, J.N. 1972. Estudio sedimentológico de la Formación Puelche en la Provincia de Buenos Aires. Revista de la Asociación Geológica Argentina 27: 5-62.
Scasso, R.A., McArthur, J.M., del Río, C., Martínez, S. y Thirlwall, M.F. 2001 - 87 SR/86 SR Late Miocene age of fossil molluscs in the “Entrerriense” of the Valdés Peninsula (Chubut, Argentina). Journal of South American Earth Sciences, 14: 319-329.
Scartascini, G. 1954. El límite Plio-Pleistoceno de la Provincia de Entre Ríos. Revista del Museo Municipal de Ciencias Naturales de Mar del Plata 1: 55-77.
Schultz, P.H., Zárate, M., Hames, W. y Camilión, C. 1998 - Impact-generated escorias as benchmarks for Argentine loess chronostratigraphy. Resúmenes del VII Congreso Argentino de Paleontología y Bioestratigrafía: 133.
Scillato-Yané, G.J. 1975b. Nuevo género de Dasypodidae (Edentata, Xenarthra) del Plioceno de Catamarca (Argentina). Algunas consideraciones filogenéticas y zoogeográficas sobre los Euphractini. Actas del I Congreso Argentino de Paleontología y Bioestratigrafía 2: 449-461, Tucumán 1974.
Scillato-Yané, G.J. 1977a. Octomylodontinae: una nueva Subfamilia de Mylodontidae (Edentata, Tardigrada).
Descripción del cráneo y mandíbula de Octomylodon robertoscagliai n. sp. procedente de la Formación Arrroyo
Chasicó (Edad Chasiquense, Plioceno temprano) del sur de la Provincia de Buenos Aires (Argentina). Algunas
consideraciones filogenéticas y sistemáticas sobre los Mylodontoidea. Publicaciones del Museo Municipal de Ciencias
Naturales. “L. Scaglia” 2: 123-140.
Scillato-Yané, G.J. 1980. Nuevo Megalonychidae (Edentata, Tardigrada) del “Mesopotamiense” (Mioceno Tardío-Plioceno) de la provincia de Entre Rios. Ameghiniana 17: 193-199.
Scillato-Yané, G.J. 1981. Nuevo Mylodontinae (Edentata, Tardigrada) del “Mesopotamiense” (Mioceno Tardío-Plioceno) de la provincia de Entre Rios. Ameghiniana 18: 29-34.
Simpson, G.G. 1940. Review of the mammal-bearing Tertiary of South America. Proceedings of the American Philosophical Society 83: 649-710.
Springer, S. y Bullis, H. R. 1960. A new species of sawshark, Pristiophorus schroederi, from the Bahamas. Bulletin of the Marine Science of the Gulf and Caribbean 10: 241-254.
Tonni, E.P., Scillato-Yané, G.J., Cione, A.L. y Carlini, A. 1998. Bioestratigrafía del Mioceno continental en el curso inferior del arroyo Chasicó, provincia de Buenos Aires. In: Resúmenes del VII Congreso Argentino de Paleontología y Bioestratigrafía, Bahía Blanca, p. 135.
Uliana, M.A. y Biddle, K.T. 1988. Mesozoic-Cenozoic paleogeographic and geodynamic evolution of southern South America. Revista Brasileira de Geociências 18: 172-190.
Woodward, A.S. 1900. On some fish-remains from the Parana Formation, Argentine Republic. Annals and Magazine of Natural History, London 6: 1-7.
Zinsmeister, W., Marshall, L.G., Drake, R. y Curtis, G. 1981 - First radioisotope (potassium-argon) age of marine Neogen Río Negro Beds in northeastern Patagonia, Argentina. Science, 212: 440.