Los roedores del “Mesopotamiense” (Mioceno tardío,
Formación Ituzaingó) de la provincia de Entre Ríos (Argentina).
Adriana Magdalena CANDELA1
1Departamento Científico Paleontología Vertebrados, Facultad de Ciencias Naturales y Museo, Paseo del Bosque s/n , 1900 La Plata, Argentina. E-mail: acandela@museo.fcnym.unlp.edu.ar
Abstract: MESOPOTAMIENSE RODENTS (LATE MIOCENE, ITUZAINGO FORMATION FO ENTRE RÍOS, PROVINCE (ARGENTINA). The Hystricognathi rodents recorded in the “Conglomerado osífero” (=“Mesopotamiense”, Late Miocene, Huayquerian Age), outcropping in the base of the Ituzaingó Formation (Entre Ríos, Argentina), show a wide species diversity, encompassing all South American superfamilies of the Suborder. Among them, the unusual assemblage of echimyids, abrocomids, neoepiblemids and Myocastor is identified, a unique composition not registered before in any other South American Cenozoic fossil fauna. In spite of the rich representativity of these “Mesopotamian” Hystricognathi, their systematics, biogeographic history, paleoenvironmental and biochronological significance remain poorly known. This contribution summarizes the state of knowledge of the Hystricognathi families from the Ituzaingó Formation (Entre Rios Province) and provides a preliminary list of species of the Suborder Hystricognathi coming from this unit, as a reference framework for future studies.
Key words: Rodentia - Hystricognathi - “Mesopotamiense” - Late Miocene - Entre Ríos Province - Argentina.
Palabras clave: Rodentia - Hystricognathi - “Mesopotamiense” - Mioceno tardío - Entre Ríos - Argentina.
Introducción
Los roedores de la Mesopotamia argentina registrados en el “Conglomerado osífero” (= “Mesopotamiense”, sensu Frenguelli, 1920; Mioceno tardío, Edad Huayqueriense; Cione et al., 2000), aflorante en la base de la Formación Ituzaingó, en la provincia de Entre Ríos, pertenecen al Suborden Hystricognathi, cuyos representantes sudamericanos son incluidos tradicionalmente dentro del Infraorden Caviomorpha. Ellos son particularmente interesantes desde un punto de vista biogeográfico y evolutivo ya que: (1) muestran una gran diversidad de especies, pertenecientes a todas las superfamilias reconocidas para el Suborden en América del Sur (Cione et al., 2000) (Apéndice 1); (2) exhiben un amplio rango de tamaño corporal, diversos diseños oclusales y cráneomandibulares, y diferentes grados de hipsodoncia (figura 1); (3) varios taxones están presentes también en otras áreas biogeográficas de Argentina y del resto de América del Sur durante gran parte del Cenozoico tardío; (4) numerosos taxones se registran sólo, con primeros y últimos registros, en esta unidad y parecen evidenciar fenómenos de extinción.
En este sentido, los Hystricognathi “mesopotamienses” ofrecen un modelo adecuado para evaluar aspectos ecológico-evolutivos y reconstruir las características paleomabientales y relaciones biogeográficas del área Mesopotámica durante el Neógeno. Estos aspectos podrán ser analizados una vez esclarecida la sistemática de la mayoría de los taxones reconocidos, los cuales necesitan una revisión sustancial (Candela, en preparación).
En esta contribución se realiza una síntesis del estado actual del conocimiento de las familias de Hystricognathi registradas en la Formación Ituzaingó, en la que se destacan aspectos sistemáticos y biogeográficos de algunos de los grupos representados. Se brinda además una lista preliminar de especies de Hystricognathi reconocidas en esta unidad, a fin de contar con un marco de referencia para futuros estudios (Apéndice 1).
Siguiendo la propuesta de Cione et al. (2000), se asume que la fauna del “Conglomerado osífero” es asignable a la Edad Huayqueriense (Mioceno tardío), pendiente de un estudio integral de correlación de las unidades fosilíferas del Neógeno de América del Sur.
Los Hystricognathi del “Mesopotamiense”
SUPERFAMILIA ERETHIZONTOIDEA
Los Erethizontidae registrados en el “Conglomerado osífero” están representados por
Paradoxomys cancrivorus Ameghino 1886, un puercoespín endémico del “Mesopotamiense” (Vucetich
y Candela, 2001). Algunos rasgos de su morfología mandibular y dentaria (figura 1.A) indican que
esta especie estaría vinculada al clado de los Erethizontinae (sensu Candela, 2004), al que pertenecen
los géneros vivientes Erethizon y Coendou, de hábitos semi y estrictamente arborícolas (Candela, 2000;
Reguero et al., en prensa). De confirmarse la pertenencia de P. cancrivorus al clado de los puercoespines
vivientes, esta especie sería el miembro más antiguo de la subfamilia, cuya historia biogeográfica
parece haberse desarrollado esencialmente en la subregión Brasílica (sensu Hershkovitz, 1958) o
región Neotropical sensu stricto (Morrone, 2001). Su presencia en el “Conglomerado osífero” representaría
además el registro más austral del clado en Argentina (Candela y Morrone, 2003).
SUPERFAMILIA OCTODONTOIDEA
Los Echimyidae son un grupo de Hystricognathi de pequeño a mediano tamaño, que incluye
numerosas especies vivientes habitantes de la subregión Brasílica (sensu Hershkovitz, 1958). Los
Echimyidae “mesopotamienses” están representados por “Eumysops” parodii Kraglievich, 1945
(Vucetich y Verzi, en Cione et al., 2000; figura 1.B), y por el endémico género y especie Haplostropha
scalabriniana Ameghino 1891. Según Vucetich y Verzi (en Cione et al., 2000) ambos taxones pertenecen
a la subfamilia Eumysopinae, un grupo que incluye formas terrestres y semisubterráneas (Reig,
1989), aunque representarían linajes diferentes de aquellos del Mioceno tardío del oeste y centro de
Argentina (Verzi et al., 1994, 1995; Vucetich, 1995; Vucetich y Verzi, 1995). Al respecto cabe señalar
que la monofila de los Eumysopinae fue recientemente cuestionada (Leite y Patton, 2002; Galewski
et al., 2005). Por lo tanto, un análisis integral de taxones extintos (e. g. Reigechimys, Pampamys,
Eumysops, Theridomysops) y vivientes (e. g. Trichomys, Clyomys, Euryzygomatomys) tradicionalmente
incluidos en los Eumysopinae (Verzi et al., 1994, 1995; Vucetich, 1995), es necesario a fin de poner a
prueba la monofilia de este grupo y afinidades de los equímidos “mesopotamienses”. Sobre esta
base se podrá examinar la historia biogeográfica de la familia, particularmente la de aquellos equímidos
del área Mesopotámica.
La presencia de “Eumysops” parodii y Haplostropha scalabriniana en el “Conglomerado osífero” sustenta la importante diversidad que los Eumysopinae tuvieron durante el Mioceno tardío, la cual habría sido mayor que la desarrollada durante el Plioceno y Pleistoceno (Vucetich y Verzi, 1995). Ambas especies, con una masa estimada de aproximadamente 1100 g., se encuentran entre los más grandes equímidos del Mioceno tardío de Argentina y entre los más pequeños Hystricognathi “mesopotamienses”, los cuales habrían convivido con los gigantes Phoberomys, Neoepiblema y Eumegamys, chinchilloideos que alcanzaron los mayores tamaños del Orden Rodentia (figura. 1. EF).
Es interesante remarcar la ausencia en el “Mesopotamiense” de los Octodontidae, una familia estrechamente emparentada con los Echimyidae, con una importante representación durante el Neógeno de Argentina (e. g. Verzi, 1999, 2001, 2002; Verzi, et al., 1999). Los Ctenomyinae, representados actualmente por el politípico género Ctenomys, se habrían diferenciando durante el Mioceno tardío (Edad Chasiquense), probablemente en el área pampásica (Verzi, 1999, 2002). Dicha diferenciación podría haber estado vinculada al deterioro climático y desarrollo de biomas abiertos inferidos para este momento (Verzi, 2002). La ausencia de Ctenomyinae en el “Mesopotamiense” (asumiendo que ésta no se debe a un error de registro), pudo haber estado condicionada por el desarrollo en el área Mesopotámica de condiciones climático-ambientales no propicias para los representantes miocénicos de la subfamilia, los cuales fueron frecuentes en Argentina Central (Verzi, 2002). Los Octondontinae se distribuyen actualmente al sur de América del Sur, principalmente en la región andina y subandina, entre los 16 y 40º de latitud sur (Mares y Ojeda, 1982). Sus representantes extintos muestran una distribución relativamente amplia en Argentina, que incluye la región pampeana, donde se registran desde el Chasiquense (Mioceno tardío) hasta el Sanandresense (Plioceno tardío; Verzi, 2001) y en el Ensenadense (Verzi et al., 2002). La ausencia de este grupo en el área Mesopotámica durante el Mioceno tardío pudo deberse también al desarrollo de condiciones ambientales no aptas para la vida de estos roedores. Sin embargo, es necesario un conocimiento mayor de la historia biogeográfica y paleobiología de los Octodontidae extintos a fin de evaluar su ausencia en el “Mesopotamiense”.
Los Myocastoridae están representados actualmente por una única especie, Myocastor coypus, distribuida en Argentina, Chile, Uruguay, Paraguay, Bolivia y sur de Brasil (Woods et al., 1992). Los registros más antiguos de este género se identifican en el “Conglomerado osífero” (Candela y Noriega, 2004) y en el “Puelchense” (Rusconi, 1945) (aceptando la correlación de al menos la base de las “capas puelchenses” con la Formación Ituzaingó; véase Cione et al., 2000). Estos registran indican que los miocastóridos “mesopotamienses” (figura 1.C) están representados sólo por Myocastor, siendo aparentemente éste el único género viviente de Hystricognathi reconocido en esta unidad y el más antiguo del Suborden identificado hasta el momento en América del Sur.
Para el “Mesopotamiense” fueron descriptas las especies endémicas Myocastor paranensis (Ameghino 1885, 1886, 1889; Rusconi, 1929) y M. obesus (Ameghino, 1889; Rusconi, 1929). Ameghino (1891) reconoció además a Colpostemma sinuata (única especie del género), también endémica del “Mesopotamiense”, aunque recientemente Candela y Noriega (2004) la refirieron al género Myocastor. Por otra parte, Rusconi (1929) asignó a “Myopotamus” priscus Gervais y Ameghino (Ensenadense - Lujanense; Ameghino, 1889: 133; Lám.V, fig. 2; Rusconi, 1929) dos fragmentos craneanos provenientes del “Mesopotamiense”, aunque está asignación no fue justificada. Las especies de Myocastor, especialmente aquellas registradas en el “Conglomerado osífero”, han sido reconocidas esencialmente por sus diferencias de tamaño, sin tener en cuenta la posibilidad de que éstas puedan deberse a variaciones ontogenéticas. La validez de las especies “mesopotamienses” de Myocastor necesita ser confirmada luego de la revisión del conjunto de las especies del género, contemplando la variación ontogenética e individual de la viviente M. coypus (Candela, en preparación).
Cabe destacar la presencia en el “Mesopotamiense” del endémico Strophostephanos jheringi, Ameghino, 1891, un roedor muy pequeño, de afinidades inciertas, que fue incluido originalmente por Ameghino (1991) en los “Echinomyidae”. Posteriormente, Mones (1986) lo consideró un género de Echimyidae, una postura seguida por McKenna y Bell (1997), quienes lo incluyeron dentro de los Myocastorinae. El status sistemático y afinidades de este roedor están siendo actualmente analizados (Candela, en preparación).
SUPERFAMILIA CHINCHILLOIDEA
Los Abrocomyidae están representados actualmente por el género Abrocoma, habitante de zonas
andinas áridas (Glanz y Anderson, 1990), y por el género Cuscomys, habitante de áreas forestadas del
norte de la cordillera peruana (Emmons, 1999). Los registros más antiguos de la familia provienen
de diferentes unidades asignadas al Mioceno tardío (Rovereto, 1914; Kraglievich, 1927; Montalvo et
al., 1996; Herbst et al., 2000). Particularmente en el “Conglomerado osífero” se reconoce a la especie
endémica Protabrocoma paranensis Kraglievich, 1927. En relación a su presencia en el “Mesopotamiense”
Vucetich y Verzi (en Cione et al., 2000: 211) señalaron que: ...“the most plausible hypothesis is that
Protabrocoma paranensis had environmental constraints different to the living species of Abrocoma. In
this sense, it is remarkable the record of a recent unusual abrocomid, showing climbing capabilities,
from the forested areas in the northern Vilcabamba cordillera (Bolivia; Cuscomys; Emmons, 1999)”.
Los abrocómidos del Mioceno tardío pudieron haber tenido un hábitat más generalizado que el de
las especies vivientes de Abrocoma. Estas últimas se habrían diferenciado más recientemente, probablemente
durante el Plioceno, en asociación a los cambios climáticos y ambientales vinculados a la
fase diastrófica Diaguita de la orogenia andina, que dio lugar a una mayor aridización de los ambientes
(Pascual et al., 1996). En este sentido, el estudio de las afinidades filogenéticas y biogeografía de
los representantes fósiles de Abrocomyidae y especialmente de Protabrocoma paranensis puede ser de
gran valor para comprender la historia evolutiva y biogeográfica de la familia, así como su significado
paleoambiental.
Los Chinchillidae “mesopotamienses” están representados por las especies Lagostomopsis pallidens (Ameghino 1886) y L. antiquus (Ameghino, 1883) (figura 1.D). Cabe destacar que un único género,
Lagostomus (Lagostomopsis), ha sido reconocido para los chinchíllidos plio-pleistocénicos (Vucetich y
Verzi, 1995). Es probable que en este género deban incluirse también los chinchíllidos del Mioceno
tardío (como aquellos reconocidos en la formaciones Arroyo Chasicó, Andalhualá, Chiquimil, Cerro
Azul, e Ituzaingó; Rovereto, 1914; Marshall y Patterson, 1981; Marshall et al., 1983; Montalvo et al.,
1996; Bondesio et al., 1980; Herbst et al., 2000). Al respecto cabe destacar que los rasgos utilizados
para diagnosticar ambos géneros corresponden esencialmente a diferencias de tamaño (Francis y
Mones, 1966; Vucetich y Verzi, 1995), las cuales son comunes entre los representantes de la especie viviente Lagostomus maximus. La identidad genérica de los chinchíllidos del Mioceno tardío necesita
ser analizada luego de una revisión sistemática de la familia que incluya los representantes extintos.
Los Dinomyidae están representados actualmente por una única especie, Dinomys branickii, habitante de zonas selváticas del este de los Andes en Brasil, Perú, Ecuador, Bolivia, Venezuela y Colombia (White y Alberico, 1992). La familia se conoce desde el Mioceno medio de Patagonia (Ameghino, 1904) y registra su mayor diversidad en el “Conglomerado osífero”, con más de 30 especies descriptas, incluidas en más de 10 géneros (e. g. Ameghino, 1885, 1886, 1891; Kraglievich, 1926, 1927, 1931, 1932; Nasif et al., 2004; Apéndice 1, figura 1.E). Sin embargo, los dinómidos en su conjunto necesitan una revisión sustancial, de hecho, varias especies “mesopotamienses” han sido fundadas originalmente en base a un único diente aislado, e. g. Pseudosigmomys paranensis Kraglievich, 1931, Pentastylomys seriei Kraglievich, 1926, “Protomegamys” coligatus Kraglievich, 1932, Teledontomys compressidens Kraglievich, 1931, Tetrastylomys castellanosi Kraglievich, 1926, Tetrastylus difussus Ameghino 1886. El registro fósil de dinómidos indica que la familia tuvo una amplia distribución geográfica en Argentina durante el Mioceno tardío, así lo demuestran aquellos taxones reconocidos en las formaciones Arroyo Chasicó, Epecuén, El Morterito, Chiquimil (Miembro El Jarillal) y Andalhualá (Rovereto, 1914; Pascual et al., 1966; Bondesio et al., 1980; Marshall y Patterson, 1981; Herbst et al., 2000). Por otra parte, en el “Conglomerado osífero” se reconocen varios géneros endémicos, e. g. Briaromys, Doellomys, Pentastylodon, Teledontomys, Pentastylomys, Pseudosigmomys (Apéndice 1).
A pesar de la rica representación de la familia en el registro fósil sudamericano (Mones, 1981; Latrubesse et al., 1997; Linares, 2004), no existe todavía un esquema filogenético que permita evaluar su historia biogeográfica. Es posible que aquellos dinómidos registrados en el oeste, centro y este de Argentina durante el lapso Chasiquense-Huayqueriense, (e. g. Tetrastylus, Potamarchus) hayan tenido una amplia distribución en momentos previos, más cercanos a la transgresión del mar “paranense” (Mioceno medio?). Este mar pudo haber fragmentado el área de distribución de estos taxones, promoviendo fenómenos de vicarianza. De cualquier modo la historia biogeográfica de la familia podrá ser evaluada luego de un estudio sistemático, especialmente de aquellos taxones provenientes de las unidades fosilíferas antes mencionadas (Nasif en preparación).
Los Neoepiblemidae representan la única familia extinta de Hystricognathi registrada en el “Conglomerado osífero”. Conocidos desde el Oligoceno tardío, alcanzaron su mayor diversidad en el “Mesopotamiense” (Apéndice 1), aunque es posible que esta diversidad sea menor a la reconocida. Los neopiblémidos se destacan por haber alcanzado el mayor tamaño corporal entre los Rodentia, una masa de aproximadamente 700 kg ha sido estimada para Phoberomys pattersoni (Sánchez Villagra et al., 2003). Se caracterizan además por presentar molariformes euhipsodontes, con diseños laminares y lofos/ídos separados por una gruesa capa de cemento interlaminar (Bondesio et al., 1975; Mones, 1981; figura 1.F). Desde que Kraglievich (1926) reconociera a los Neoepiblemidae, la composición taxonómica y diagnosis del grupo han variado considerablemente (Bondesio et al., 1975; Mones, 1981; Bondesio y Bocquentin-Villanueva, 1988; Bocquentin-Villanueva et al., 1990; Sant´anna-Filho, 1994). Kraglievich (1926) consideró como Neoepiblemidae a Neoepiblema y Dabbenea, y vinculó a la familia más estrechamente con los Chinchillidae (Kraglievich, 1940). Esta posición fue seguida por Bondesio et al. (1975) quienes además incluyeron entre los neopiblémidos a Perimys, un género del Mioceno de Patagonia (Kramarz, 2002). El status sistemático de Perimys scalabrinianus Ameghino 1889, la única especie del género identificada en el “Conglomerado osífero”, no pudo ser confirmado, dado a que el material tipo y único ejemplar conocido no ha sido hallado luego de su descripción original. Bondesio y Bocquentin-Villanueva (1988) consideraron a Phoberomys (previamente incluido en los Dinomyidae) un género válido de Neoepiblemidae, y Negri y Ferigolo (1999) a Phoberomys(incluyendo a Dabbenea), Neoepiblema (Ameghino 1889) (incluyendo a Euphilus) y Eusigmomys Ameghino (1904). La inclusión de Scotamys en los Neoepiblemidae extiende el registro de la familia hasta el Oligoceno tardío (Kramarz, 2001). Un estudio preliminar de los neopiblémidos “mesopotamienses” indica que ellos comparten varios rasgos con los Eumegamyinae (un grupo perteneciente a los Dinomyidae; Mones, 1981, Candela; en preparación).
La presencia de Neoepiblemidae en el “Mesopotamiense”, Urumaco (Mioceno tardío, Venezuela; Mones, 1981; Bondesio y Bocquentin-Villanueva, 1988), y Acre (Mioceno tardío, Brasil; Sant´Anna Filho, 1994; Bocquentin-Villanueva et al., 1990; Negri y Ferigolo, 1999) sugiere una vinculación biogeográfica del área de estudio con la subregión Brasílica (sensu Hershkovitz, 1958). Ésta familia representa además uno de los linajes que desde un punto de vista faunístico permite distinguir el área Mesopotámica del noroeste y centro de Argentina durante el Mioceno tardío, tal como lo indican otros grupos de mamíferos (xenartros y ungulados, véase Cione et al., 2000; Candela y Morrone, 2003).
SUPERFAMILIA CAVIOIDEA
Los Hydrochoeridae están representados en la actualidad por Hydrochaeris hydrochaeris, distribuida
en gran parte de América del Sur (Eisenberg y Redford, 1999). Esta especie es el roedor viviente de
mayor tamaño y el único sobreviviente de la familia. Los hidroquéridos se registran desde el
Chasiquense, y tuvieron una diversidad importante durante el Mioceno tardío y Plioceno (Mones,
1991). En el “Mesopotamiense” han sido reconocidos numerosos géneros y especies, aunque es posible que esta diversidad sea menor. De hecho, un estudio en preparación indica que varias especies
de carpinchos “mesopotamiense” serían sinónimas (Vucetich et. al., en prensa). Por este motivo,
sólo se brinda aquí una lista de los géneros representados en esta unidad (Apéndice 1).
Los cávidos “mesopotamienses” están representados también por numerosos géneros y especies, aunque, como en el caso de los Hydrochoeridae, ellos necesitan una revisión sustancial. Por lo tanto, sólo se brinda una lista de aquellos géneros previamente reconocidos en esta unidad (Cione et al., 2000) (Apéndice 1).
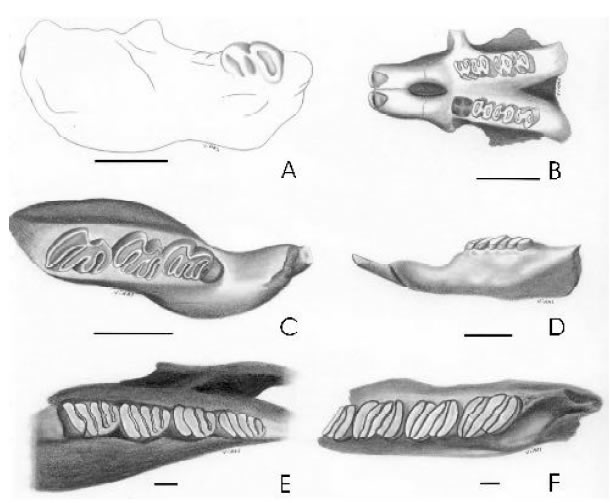
Fig. 1. Roedores de la Formación Ituzaingó. A, Paradoxomys cancrivorus, fragmento mandibular derecho con el incisivo y el m2, vista lingual (modificado de Vucetich y Candela, 2001); B,“Eumysops” parodii, fragmento de cráneo con P4-M3 izquierdos y M1-M3 derechos, vista oclusal (modificado de Kraglievich, 1945); C, Myocastor sp., fragmento mandibular izquierdo con dp4-m2, vista oclusal (modificado de Candela y Noriega, 2004); D, Lagostomopsis antiquus, fragmento mandibular izquierdo con el incisivo y la serie p4-m2, vista labial (modificada de Ameghino, 1889: Lám. XXVI, fig. 9a); E, Eumegamys paranensis, fragmento mandibular derecho con p4-m3, vista oclusal (modificado de Kraglievich, 1926); F, Phoberomys burmeisteri, fragmento mandibular izquierdo con parte del p4 y la serie m1-3, vista oclusal (modificado de Kraglievich, 1926). Escala: 1 cm.
Discusión
De la información precedente se destaca la importante diversidad de los Hystricognathi registrados en la Formación Ituzaingó, así como la necesidad de esclarecer la sistemática de la mayoría de los taxones reconocidos en esta unidad. Sobre esta base se podrá analizar su historia biogeográfica y precisar su significado biocronológico, a fin de lograr una más ajustada correlación temporal del “Mesopotamiense” con otras unidades tradicionalmente asignadas al Mioceno tardío y Plioceno de América del Sur.
La presencia compartida de ciertos géneros, e. g. Lagostomopsis, Tetrastylus, Potamarchus, Cardiomys, en la Formación Ituzaingó y en otras unidades de Argentina asignadas al Mioceno tardío (e. g. Formación Arroyo Chasicó, Formación Chiquimil, Formación Andalhualá, Formación Cerro Azul, Formación Epecuén) podría ser el resultado de una amplia y más antigua distribución de estos taxones, al menos desde la Edad Chasiquense.
Los neopiblémidos y el puercoespín Pardoxomys cancrivorus, una especie vinculada a los Erethizontinae (sensu Candela, 2004), expresaría una mayor relación del área Mesopotámica con la subregión Brasílica (senus Hershkovitz, 1958) y sustenta la hipótesis de su diferenciación de las restantes áreas de Argentina, al menos desde el Mioceno tardío (Cione et al., 2000; Candela y Morrone, 2003).
Los roedores del “Mesopotamiense” exhiben el rango de tamaño corporal más amplio registrado entre los Hystricognathi del Cenozoico de Argentina. Entre ellos se identifican además taxones braquiodontes (e. g. puercoespines), protohipsodontes (e. g. Myocastor) y euhipsodontes (e.g. neopiblémidos, hidroquéridos), con diseños oclusales complejos, como en estos últimos, y simplificados, como el de los chinchíllidos (figura 1). Esta diversidad, que parece indicar el desarrollo de diversos hábitos y hábitats, necesita ser estudiada desde un punto de vista eco-morfológico, poniendo a prueba la existencia de “constraints” filogenéticos en los caracteres analizados, a fin de evaluar su significado adaptativo y paleoambiental.
Por otra parte, entre los Hystricognathi “mesopotamienses” se registra la inusual asociación de equímidos, abrocómidos, neopiblémidos y Myocastor, una composición única de taxones, hasta ahora no reconocida en ninguna de las faunas fósiles del Cenozoico de América del Sur, cuyo significado ecológico-evolutivo y biogeográfico debe ser aún analizado.
Agradecimientos
Deseo expresar mi agradecimiento a F. Aceñolaza por invitarme a participar de este trabajo, a A. Kramarz por facilitarme el acceso de las colecciones del Museo Nacional de Ciencias Naturales “Bernardino Rivadavia”, y a J. Noriega por brindarme valiosa información acerca de la problemática del “Mesopotamiense”.
Este trabajo es una contribución los PICT 11928 y 8395 (AGENCIA).
Apéndice 1
Familia Erethizontidae
Paradoxomys Ameghino, 1885 nomen dubium
P. cancrivorus Ameghino, 1886 nomen dubium
Familia Echimyidae
Haplostropha Ameghino, 1891
H. sacabriniana Ameghino, 1891
“Eumysops” parodii Kraglievich, 1945
Familia Myocastoridae (subfamilia Myocastorinae sensu McKenna y Bell, 1997)
Myocastor Kerr, 1792
M. paranensis (Ameghino, 1885)
M. obesus (Ameghino, 1889)
M. sinuata (Ameghino, 1991)
Myocastoridae?
Strophostephanos Ameghino, 1991
S. jheringi Ameghino, 1991
Familia Abrocomyidae
Protabrocoma Kraglievich, 1927
P. paranensis Kraglievich, 1927
Familia Chinchillidae
Lagostomopsis Kraglievich, 1926
L. pallidens (Ameghino, 1889)
L. antiquus (Ameghino, 1883)
Familia Dinomyidae
Briaromys Ameghino, 1889.
B. trouessartianus Ameghino, 1889
“Discolomys” Ameghino, 1889
“D”. cuneus Ameghino, 1889
Doellomys Alvarez, 1947
D. parcus Alvarez, 1947
Eumegamys Kraglievich, 1926 (= Megamys, Isostylomys, Carlesia, Diaphoromys, Rusconia, Protomegamys,
sensu McKenna y Bell, 1997).
E. contortus (Ameghino, 1889)
E. scalabrinianus Kraglievich, 1926
E. paranensis Kraglievich, 1926
E. dubius Kraglievich, 1932
“E”. depressidens (Ameghino, 1885)
Carlesia Kraglievich, 1926
C. pendolai Kraglievich, 1926
Diaphoromys Kraglievich, 1931
D. compressidens Alvarez, 1947
D. gamayensis Kraglievich, 1931
D. mesopotamicus Kraglievich, 1932 (= D. artussi Kraglievich, 1934)
Isostylomys Kraglievich, 1926
I. ameghinoi Kraglievich, 1932
I. laurillardi (Ameghino, 1883)
Protomegamys Kraglievich, 1932
P. coligatus Kraglievich, 1932
Rusconia Kraglievich, 1931
R. crassidens Kraglievich, 1931
Eumegamysops Kraglievich en Alvarez, 1947
E. praependens (Ameghino, 1886)
Gyriabrus Ameghino, 1891
G. glutinatus Ameghino, 1891
G. holmbergi (Ameghino, 1885)
G. indivisus Kraglievich, 1932
G. rebagliattii Kraglievich, 1932
Paranamys Kraglievich, 1934 (=Olenopsis Ameghino 1889)
O. typicus, (Ameghino, 1889)
Pentastylodon Kraglievich en Alvarez, 1947
P. racedi (Ameghino, 1885)
Pentastylomys Kraglievich, 1926
P. seriei Kraglievich, 1926
Potamarchus Burmeister, 1885
P. murinus Burmeister, 1885
P. sigmodon Ameghino, 1891
Pseudosigmomys Kraglievich, 1931
P. paranensis Kraglievich, 1931
Telodontomys Kraglievich, 1931
T. compressidens Kraglievich, 1931
Tetrastylomys Kraglievich, 1926
Tetrastylomys castellanosi Kraglievich, 1926
Tetrastylus Ameghino, 1886 (=Loxomylus, Loxopygus, Tetrastylopsis, Protelicomys, sensu McKenna y
Bell, 1997).
T. aguilari Kraglievich, 1932
T. diffusus Ameghino, 1886
T. laevigatus (Amgehino, 1885)
T. robustus Kraglievich, 1932
T. (Protelicomys) atavus Kraglievich, 1931
Familia Neoepiblemidae
Neoepiblema Ameghino, 1889 (= Euphilus Ameghino, 1889; Epiblema Ameghino, 1886, sensuMcKenna y Bell, 1997).
N. horridula Ameghino, 1886
N. ambrosettianus (Ameghino, 1889)
Phoberomys Kraglievich, 1926 (= Dabbenea Kraglievich, 1926)
P. burmeisteri (Ameghino, 1891) Kraglievich, 1926
P. praecursor Kraglievich, 1932
P. insolita (Kragievich, 1940)
P. lozanoi (Kragievich, 1940)
P. minima (Kragievich, 1940)
Perimys Amghino, 1887
P. scalabrinianus Ameghino, 1889
Familia Hydrochoeridae
Anatochoerus Mones y Vucetich, 1991
Anchimysops Kraglievich, 1927
Cardiatherium Ameghino, 1883
Cantracavia Burmeister, 1885
Kiyutherium Francis y Mones, 1965
Plexochoerus Ameghino, 1886
Procardiatherium Ameghino, 1885
Protohydrochoerus Rovereto, 1914
Familia Caviidae
Cardiomys Ameghino, 1885
Caviodon Ameghino, 1885
Paleocavia Ameghino, 1889
Parodimys Kraglievich, 1932
Pliodolichotis Kraglievich, 1927
Prodolichotis Kraglievich, 1932
Apéndice 1. Lista preliminar de especies de Hystricognathi “mesopotamienses” (la sistemática de las familias Neoepiblemidae y Dinomyidae sigue parcialmente las propuestas de Mones, 1986; McKenna y Bell, 1997 y Negri y Ferigolo, 1999). Los cavioideos sólo son citados a nivel genérico (véase el texto).
Bibliografía
Alvarez, E. F. de. 1947. Nota preliminar sobre nuevos roedores “Eumegamyidae”. Revista de la Asociación Geológica Argentina 2 (1): 59-64.
Ameghino, F. 1983. Sobre una nueva colección de mamíferos fósiles recogidos por el Profesor Scalabrini en las barrancas del Paraná. Boletín de la Academia Nacional de Ciencias de Córdoba 5: 112-113.
Ameghino, F. 1885. Nuevos restos de mamíferos fósiles oligocenos recogidos por el profesor Pedro Scalabrini y pertenecientes al Museo Provincial de la Ciudad de Paraná. Boletín de la Academia Nacional de Ciencias de Córdoba 8: 5-207.
Ameghino, F. 1886. Contribución al conocimiento de los mamíferos fósiles de los terrenos terciarios antiguos de Paraná. Boletín de la Academia Nacional de Ciencias de Córdoba 9: 5-228.
Ameghino, F. 1887. Enumeración sistemática de las especies de mamíferos fósiles colectados por Carlos Ameghino en los terrenos eocenos de Patagonia austral y depositados en el Museo de La Plata. Boletín del Museo de La Plata 1: 1-26 (del apartado).
Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Boletín de la Academia Nacional de Ciencias de Córdoba 6: xxxii + 1-127.
Ameghino, F. 1904. Nuevas especies de mamíferos cretácicos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 57: 162-175, 327-341.
Bocquentin-Villanueva, J. Souza-Filho, J. P. y Negri, F. R. 1990. Neoepiblema acreensis, sp. n. (Mammalia, Rodentia) do Neógeno de Acre, Brasil. Boletim do Museu Paraense Paraná Emilio Goeldi 2: 65-72.
Bondesio, P., Pascual, R. y Vucetich, M. G. 1975. Los Neoepiblemidae (Rodentia, Caviomorpha): su caracterización y sus relaciones filogenéticas con los Chinchillidae. Actas del primer Congreso Argentino de Paleontología y Bioestratigrafía, Tomo II: 431-447. Tucumán.
Bondesio, P. y Bocquentin-Villanueva, J. 1988. Novedosos estos de Neoepiblemidae (Rodentia, Hystricognathi) del Mioceno tardío de Venezuela. Inferencias paleoambientales. Ameghinana 25(1): 31-37.
Candela, A. M. 2000. [Los Erethizontidae (Rodentia, Hystricognathi) fósiles de Argentina. Sistemática e historia evolutiva y biogeográfica. Tesis Doctoral, Facultad de Ciencias Naturales y Museo de La Plata, 352 p. Inédita].
Candela, A. M. y Noriega, J. I. 2004. Los coipos (Rodentia, Caviomorpha, Myocastoridae) del “Mesopotamiense” (Mioceno tardío; Formación Ituzaingó) de la provincia de Entre Ríos, Argentina. En: F. G. Aceñolaza (Ed.), Temas de la Biodiversidad del Litoral Fluvial Argentino, serie INSUGEO, Miscelánea, 12:5-12.
Candela, A. M. 2004. A new giant porcupine (Rodentia, Erethizontidae) from the late Miocene of northwestern Argentina. Journal of Vertebrate Paleontology 24 (3): 732-741.
Candela, A. M. y Morrone, J. J. 2003. Biogeografía de puercoespines neotropicales (Rodentia, Hystricognathi): Integrando datos fósiles y actuales a través de un enfoque panbiogeográfico. Ameghiniana 40 (3): 361-368.
Cione, A. L., M. Azpelicueta, M. Bond, A. A. Carlini, J. R. Casciotta, M. A. Cozzuol, M. de la Fuente, Z. Gasparini, F. J. Goin, J. Noriega, G. J. Scillato-Yané, L. Soilbelzon, E. P. Tonni, D. Verzi, M. G. Vucetich. 2000. Miocene vertebrates from Entre Ríos province, eastern Argentina. En: F. G. Aceñolaza y R. Herbst (eds.), El Mio-neógeno Argentino. Insugeo. Serie Correlación Geológica 14: 191-237.
Emmons, L. H. A new genus and species of Abrocomid rodents from Perú (Rodentia: Abrocomidae). American Museum Novitates 3279: 1-14.
Francis J. C. y Mones, A. 1966. La presencia de vizcachas [(Lagostomys (Lagostomopsis) spicatus Amegh,)] en la Formación Kiyú, Dpto. de San José R. O del Uruguay. Kraglieviana 22: 147-168.
Frenguelli, J. 1920. Contribución al conocimiento de la geología de Entre Ríos. Boletín de la Academia Nacional de Ciencias de Córdoba 24: 55-256.
Galewski, T., Mauffrey J. F., Leite, Y. L. R., Patton, J. L., Douzery, E. J. P., 2005. Ecomorphological diversification among South American spiny rats (Rodentia; Echimyidae): a Phylogenetic and chronological approach. Molecular Phylogenetics and Evolution 34: 601-6015.
Glanz, W. E. y Anderson, S. 1990. Notes on Bolivian Mammals. 7. A new species of Abrocoma (Rodentia) and the relationships of the Abrocomidae. American Museum Novitates 291: 1-32.
Herbst, R., Anzótegui, L. M., Esteban, G., Mautino, L. R., Morton, S. y Nasif, N. Síntesis paleontológica del Mioceno de los valles Calchaquíes, noroeste argentino. Miocene vertebrates from Entre Ríos province, eastern Argentina. En: F. G. Aceñolaza, y R. Herbst (eds.), El Mio-neógeno Argentino. Insugeo. Serie Correlación Geológica 14: 263-268.
Hershkovitz, P. 1958. A geographic classification of Neotropical mammals. Fieldiana (Zoology) 36 (6): 581-620.
Kraglievich, L. 1926. Los grandes roedores Terciarios de la Argentina y sus relaciones con ciertos géneros pleistocenos de las antillas. Anales del Museo Nacional de Historia Natural “Bernardino Rivadavia”, Tomo XXXIV:122-135.
Kraglievich, L. 1927. Nota preliminar sobre nuevos géneros y especies de la fauna argentina. Physis 8: 591-598.
Kraglievich, L. 1931. Nuevos géneros de roedores eumegámidos. Physis 10: 392-39.
Kraglievich, L. 1932. Diagnosis de nuevos géneros y especies de roedores cávidos y eumegámidos fósiles de la Argentina. Rectificación genérica de algunas especies conocidas, y adiciones al conocimiento de otras. Anales de la Sociedad Científica Argentina 115: 155-181 y 211-237.
Kraglievich, L. 1940. Descripción detallada de diversos roedores argentinos terciarios clasificados por el autor. En: obras Completas y Trabajos Científicos Inéditos de Lucas Kraglievich. A. J. Torcelli, Compilador. Obras de Geología y Paleontología, 2: 297-330.
Kramarz, 2001: Revision of the family Cephalomyidae (Rodentia, Caviomorpha) and new cephalomyids from the Early Miocene of Patagonia. Palaeovertebrata 30 (1-2): 51-88).
Kramarz, A. 2002: Roedores chinchilloideos (Hystricognathi) de la Formación Pinturas, Mioceno temprano – medio de la provincia de Santa Cruz, Argentina. Revista del Museo Argentino de Ciencias Naturales 4 (2): 167-180.
Latrubesse, E. M., Bocquentin, J., Santos, J. C. R., Ramonell, C. G. 1997. Paleoenvironmental model for the Late Cenozoic of Southwestern Amazonia: Paleontology and Geology. Acta Amazonica 27 (2): 103-118.
Leite, Y. L. R. y Patton, J. L. 2002. Evolution of South American spiny rats (Rodentia, Echimyidae): the starphylogeny hypothesis revisited. Molecular Phylogenetics and Evolution 24: 455-464.
Linares, O. J. 2004. Bioestratigrafía de la fauna de mamíferos de las formaciones Socorro, Urumaco y Codore (Mioceno medio-Plioceno temprano) de la región de Urumaco, Falcon, Venezuela. Paleobiología Neotropical 1:1-26.
Mares, M. A. y Ojeda, R. A. 1982. Patterns of diversity and adaptation in South American Hystricognath rodents. En: M. A. Mares y H. H. Genoways (eds.), Mammalian biology in South America Special Publication Series, Pymatuning Laboratory of Ecology, University of Pittsburgh, Pennsylvania 6: 393-432.
McKenna, M. C. y Bell, S. K. 1997. Classification of Mammals Above the Species Level. Columbia University Press, New York, 631 pp.
Marshall, L. y Patterson, B. 1981. Geology and geochronology of the mammal-bearing Tertiary of the Valle de Santa María and Río Corral Quemado, Catamarca Province, Argentina. Fieldiana Geology 9:1-80.
Marshall, L. G., Hoffstetter, R. y Pascual, R., 1983. Mammals and stratigraphy. Geochronology of the continental mammal-bearing Tertiary of South America. Palaeovertebrata, Mémoire Extraordinare, 1-93.
Mones, A. 1981. Sinopsis sistemática preliminar de la familia Dinomyidae (Mammalia: Rodentia: Caviomorpha). Anais do II Congresso Latino-Americano de Paleontología II: 605-619.
Mones, A. 1986. Paleovertebrata Sudamericana. Catálogo sistemático de los Vertebrados fósiles de América del Sur. Parte I. Lista preliminar y Bibliografía. Courier Forschunginstitut Senckenberg 82: 1-625.
Mones, A. 1991. Monografía de la familia Hydrochoeridae (Mammalia: Rodentia). Courier Forschungsinstittut Senckenberg 134: 1-235.
Montalvo, C. I., Cardonatto, M. C., Visconti, G., Verzi, D. H. y Vucetich. M. G. 1996. Vertebrados de la Formación Cerro Azul (Mioceno tardío) del valle de Quehué, provincia de la Pampa, Argentina. Actas VI Jornadas pampeanas de Ciencias Naturales, Coprocna: 159-165.
Morrone, J. J. 2001. Biogeografía de América Latina y el Caribe. Monografías de la Sociedad Entomológica Aragonesa, nro. 3, Zaragoza (España).
Nasif, N. L., Aceñolaza F. G., y Esteban G. I., 2004. Nuevo material de xenartros y roedores para la Formación ituzaingó (Mioceno superior), provincia de Entre Ríos, Argentina. Reunión Anual de Comunicaciones Asociación Paleontológica Argentina, Resúmenes: 20. Diamante.
Negri, F. R. y Ferigolo, J. 1999. Anatomia craniana de Neoepiblema ambrosettianus (Ameghino, 1889) (Rodentia, Caviomorpha, Neoepiblemidae) do Mioceno Superior-Plioceno, Estado do Acre, Brasil, e revisâo das espêcies do gêneo. Boletim do Museu Paraense Paraná Emilio Goeldi Série Ciências da Terra Vol. II: 3-80.
Pascual, R. Ortega Hinojosa, E. J. Gondar, D. y Tonni, E. P. 1966. Paleontografía Bonaerense, Fasc. IV Vertebrata. A. V. Borrello (Ed.). Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, 202 pp.
Pascual, R., Ortiz Jaureguizar, E. y Prado, J. L. 1996. Land Mammals: Paradigm for Cenozoic South American Geobiotic Evolution. En: G. Arratia (Ed.), Contribution of Southern South America to Vertebrate Paloentology. Munchner Geowissenschaftliche Abhandlungen 30: 265-319.
Eisenberg, J. F y Redford, K. H. 1999. Mammals of the Neotropics. The Central Neotropics. Vol 3. Ecuador, Perú, Bolivia, Brasil. The University of Chicago Press.
Reguero, M. A., Candela, A. M. y Alonso, R. En prensa. Biochronology and biostratigraphy of the Uquía Formation (Pliocene-Early Pleistocene, NW of Argentina) and its significance in the Great American Biotic Interchange. Journal of South American Earth Sciences.
Reig, O. 1989. Karyotypic repatterning as one triggering factor in cases of explosive Speciation. En: A. Fontdevila (Ed.), Evolutionary Biology of Transient Unstable Populations. Springer-Verlag. Berlin: 246-289.
Rovereto, C. 1914. Los estratos araucanos y sus fósiles. Anales del Museo Nacional de Historia Natural de Buenos Aires 25: 1-247.
Rusconi, C. 1929. Revisión de las especies fósiles argentinas del género Myocastor, con descripción de nuevas especies. Anales de la Sociedad Argentina de Estudios Geográficos “G.E.A”. Tomo III, Nº 2: 504-518.
Rusconi, 1945. Un nuevo roedor del Entrerriano: Eumysops parodii, n. sp. Anales de la sociedad Científica Argentina, CXL: 16-21.
Sánchez Villagra, M., Aguilera, O. y Horovitz, I. 2003. The anatomy of the World´s largest extinct rodent. Science: 1708-1710.
Verzi, D. H., Vucetich, M. G. y Montalvo, C. 1994. Octodontid-like Echimyhidae (Rodentia); Un Upper Miocene episode in the radiation of the Family. Palaeovertebrata 23 (1-4) 199-210.
Verzi, D. H., Vucetich, M. G. y Montalvo, C. 1995. Un Nuevo Eumysopinae (Rodentia, Echimyidae) del Mioceno tardío de la provincia de La Pampa y consideraciones sobre la historia de la subfamilia. Ameghiniana 32: 191-195.
Verzi, D. H., Tonni, E. P., Scaglia, O. A. y San Cristóbal, J. O. 2002. The fossil record of the desert-adapted South American rodent Tympanoctomys (Rodentia, Octodontidae). Paleoenvironmental and biogeographic significance. Palaeogeography, Palaeoclimatology, Palaeoecology 179: 149-158.
Verzi, D. H. 1999. The dental evidence on the differentiation of the Ctenomyinae rodents (Caviomorpha, Octodontidae, Ctenomyinae). Acta Theriologica 44 (3): 263-282.
Verzi, D. H. 2001. Phylogenetic position of Abalosia and the evolution of the extant Octodontinae (Rodentia, Caviomorpha, Octodontidae). Acta Theriologica 46(3): 243-268.
Verzi, D. H. 2002. Patrones de evolución morfológica en Ctenomyinae (Rodentia, Octodontidae). Mastozoología Neotropical 9: 309-328.
Vucethich, M. G. 1995. Theridomysops parvulus (Rovereto, 1914), un primitivo Eumysopinae (Rodentia, Echimyidae) del Mioceno tardío de Argentina. Mastozoología Neotropical 2: 167-172.
Vucetich M. G. y Verzi, D. H. 1995. Los roedores caviomorfos. En: M. T. Alberdi. G. Leone y E. P. Tonni (eds.). Evolución Biológica y Climática de la Región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental. Monografías Museo Nacional de Ciencias Naturales de Madrid, 12: 213-225.
Vucetich, M. G. y Candela, A. M. 2001. Paradoxomys cancrivorus Ameghino, 1885 (Rodentia, Hystricognathi, Erethizontidae): the first porcupine from the “Mesopotamiense” (late Miocene). Ameghiniana 38: 147-150.
Vucetich, M. G., Dechamps, C., Olivares, A. y Dozo, T. 2004. Capybarras size, shape and time: a model kit. Acta Paleontológica Polonica. En Prensa.
Vucetich, M. G., Verzi, D. H. y Hartenberger, J. L. 1999. Review and analysis of the radiation of the South American Hystricognathi (Mammalia, Rodentia). Comptes Rendus de l’ Académie des Sciences, Paris 392: 763-769.
White, T. G. y Albertico, M. S. 1992. Dinomys branickii. Mammalian species 410: 1-4.
Woods, C. A., Contreras, L., Willner-Chapman, G., Whidden, H.P. 1992. Myocastor coypus. Mammalian Species 398: 1-8.