Crustáceos Decápodos del Litoral Fluvial Argentino
Pablo A. COLLINS1, Verónica WILLINER1 y Federico GIRI1
1Instituto Nacional de Limnología (CONICET-UNL), José Maciá 1933, 3016 Santo Tomé, Santa Fe, Argentina email: pcollins@arnet.com.ar
Abstract: Decapod Crustaceans of Argentinean Littoral Fluvial. The decapods is a group of crustaceans that has not been deeply studied in this freshwater environment, although during the last years there are some more papers making reference to these animals. In this work, It is showed a list of the Superior Crustaceans Superorden Eucarida that inhabits Argentinean territory. The data is based on some observations carried out by this work group and bibliography. Results and observations done on the biodiversity, trophic ecology, distribution, activity, growth, behaviour, ecotoxicology show it is developed as well as other work lines carried out by other investigators. This wide range of information has given enough material to argue about the importance of this group in the Argentinean Littoral Fluvial and even more to make people aware of the need for an increase in the searching.
Key words: decapods, biodiversity, crabs
Palabras clave: decápodos, biodiversidad, camarones, cangrejos, Trichodactylidae, Palaemonidae, Aeglidae, Sergestidae, ParastacidaeIntroducción
El orden Decapoda integra junto con otros dos ordenes (Amphionidacea y Euphasiaceae) el Superorden Eucarida Calman, 1904 del Subphylum Crustacea Brünnich, 1772. Los Eucáridos son un grupo muy diversificado en el que se han agrupado las taxas con un caparazón completo formando un cefalotórax compuesto por los segmentos cefálicos y torácicos.
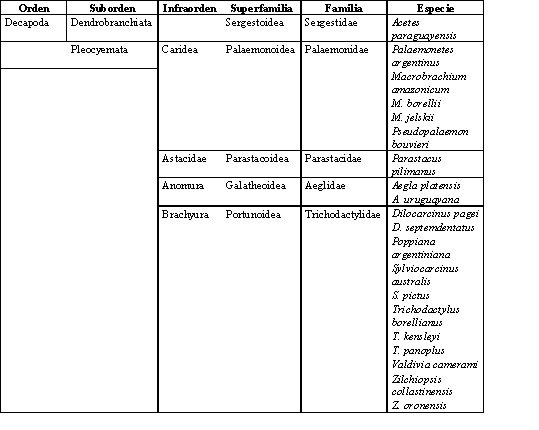
La unidad taxonómica Decapoda registra más de 8500 especies, en su mayoría restringida a áreas marinas (Brusca & Brusca, 1990). Las especies de ambientes dulciacuícolas en América del Sur están agrupadas en siete familias (Manning & Hobbs, 1977; Boschi, 1981; Rodriguez, 1981, 1992; Magalhães & Türkay, 1996 a, b, c; Morrone & Lopretto, 2001). Sin embargo, el número en los ambientes del litoral fluvial Argentino disminuye a cinco familias hasta el momento, sistemáticamente ordenadas según Martin & Davis (2001) en Sergestidae, Palaemonidae, Parastacidae, Aeglidae y Trichodactylidae (Tabla 1). Muchos de los integrantes a nivel de especie, género y en algunos casos familia son endémicos de esta región.
Los representantes de estas unidades taxonómicas habitan en bañados, esteros, lagunas, riberas y cauces principales de los ríos del sistema del Plata. Algunos construyen cuevas, otros nadan abiertamente en la columna de agua o se posan entre la vegetación y/o algún otro substrato como puede ser el sedimento de fondo.
Diversidad de decápodos
En la última década se han hallado especies de camarones y cangrejos aún no registradas en la región del río Paraná y sus afluentes, aumentando entre un 30 y 50 % el número de especies reconocidas para Argentina en la década del 80'. Esto da un indicio de la necesidad de proseguir con muestreos exploratorios sistemáticos ya que el número de especies presentes y no muestreadas puede ser aun mayor.
Entre los ejemplares conocidos como camarones se identifican seis especies pertenecientes a las familias Sergestidae y Palaemonidae (Boschi, 1981; Pettovello, 1996; Collins, 2000 a). Mientras que solo una especie de langosta (Parastacidae) esta presente en arroyos de la Mesopotámia Argentina y dos especies de cangrejos pancoras correspondientes a la familia Aeglidae. Los cangrejos tricodactílidos son los más diversos del Sistema del Plata reconociéndose diez especies (Tabla 1) (Magalhães & Türkay, 1996 a, b,c; Morrone & Lopretto, 2001; Collins et al ., 2002).
La cantidad de especies hasta hoy registrada puede verse modificada por migraciones y eventos continuos de especiación. Diferencias intraespecíficas importantes en la morfología de poblaciones aisladas geográficamente fueron observadas mediante análisis de morfometría geométrica en algunas especies de Aegla (Giri & Collins, en prensa a). Estas distancias morfológicas nos podrían indicar el efecto de alguna causa ecológica y/o diferencias evolutivas en poblaciones aisladas.
La fauna de estos crustáceos se considera proveniente de una ingresión de especies del Atlántico a través del río de la Plata, otras del Amazonas a través de conexiones efímeras con el río Paraguay o afluentes de este y otras del río Alto Paraná con ejemplares originarios de la región basilera de la “Mata Altantica” (Schmitt, 1942 ; Ringuelet, 1949 a; b; Lopretto, 1980; Collins et al ., 2002).
A continuación se mencionan algunos aspectos biológicos y ecológicos de este grupo quedando muchos temas aún sin tratar para poder comprender el funcionamiento del grupo en los sistemas acuáticos continentales.
Abundancia
La abundancia de este grupo varía con las especies, los ambientes, la presencia y tipo de vegetación acuática, la abundancia de presas, predadores y competidores, el momento del año en relación con la altura del río, la conexión de la laguna con el río y la temperatura del agua. Estos son algunos de los factores que alterarían la abundancia observando por ejemplo en P. argentinus densidades superiores a 300 ind/m 2 mientras que en otro momento del año no se registran ejemplares (Collins, 2000 b). Las diferencias temporales y espaciales en las densidades indican movimientos masivos de las poblaciones entre distintos tipos de ambientes.
Movimientos
El sistema del Plata tiene una superficie aproximada de 2,8 millones de km 2 (Bonetto & Waiss, 1995), en el cual se encuentra una gran diversidad de cuerpos acuáticos en función de su estabilidad, relación entre los cuerpos lóticos y lénticos y su composición florística y faunística.
Estos ambientes, en su mayoría, son afectados temporalmente por el ingreso y la renovación del agua de los ríos. Esto determina que los animales se encuentren en una situación de compromiso entre permanecer en un ambiente en vías de desecación junto a la posibilidad de morir, o migrar buscando un lugar con condiciones aptas para subsistir. Al disminuir el nivel de altura del agua, se pierden refugios y se concentran las poblaciones de cada especie. Esto determina un aumento en el contacto de un individuo con otro, favoreciendo las interrelaciones intraespecíficas e interespecíficas. Por otra parte, las comunidades que serían integrantes de su espectro trófico natural se concentran en las zonas aún con agua (Bonetto & Waiss, 1995; Williner & Collins, 2002 a, b). Aunque al mismo tiempo podría aumentar la competencia por espacio y alimento, como también incrementar el riesgo a ser depredados por peces, reptiles, aves y mamíferos al diminuir el nivel del agua y estar más expuestos. Todos estos elementos actuarían de manera conjunta, afectando la supervivencia e induciendo algún tipo de movimiento poblacional (Fernandez & Collins, en prensa).
Además, se han registrado movimientos entre zonas vegetadas, libres de vegetación, fondo y columna de agua en camarones y cangrejos según un ritmo diario (Renzulli & Collins, 2001; Williner & Collins, 2002 a, b)
Ecología trófica
La ecología trófica de los decápodos obliga a una interpretación integral del proceso de alimentación, el cual implica definir el tiempo y modo de localización, captura, manipulación y finalmente consumo de las presas con su asimilación (Giri et al ., 2002).
En Dilocarcinus pagei se ha observado que el alimento es tanteado y tomado con una o ambas quelas, luego es conducido hacia el tercer maxilípedo con la ayuda de los periópodos. En camarones la captura puede ser más activa llegando a perseguir a sus presas en la columna de agua, luego el manipuleo es similar a los cangrejos (Giri et al ., 2002).
Los géneros Macrobrachium y Palaemonetes tienen un estómago sencillo, compuesto por una cámara cardíaca y otra llamada pilórica. Esta última tiene paredes delgadas formando una bolsa con gran cantidad de repliegues. No se observan estructuras quitinosas o calcáreas como en otros camarones. La cámara cardíaca de M. borelli tiene en su pared anterior elementos de sostén levemente esclerosados y en la superficie ventral de la cámara pilórica un dispositivo triangular con bordes endurecidos cubierto por sedas de diferente longitud. Estas sedas estarían cumpliendo una función de transporte del material ingerido hacia el intestino (Boschi, 1981; Collins, 2000 b). Entre las especies varían el número, forma, tamaño, grado de sutura y calcificación. Así en el grupo de los Dendrobranchiata, representado en aguas continentales por el género Acetes , se observa cierta uniformidad y poco desarrollo en la armadura del estómago. Esta característica junto a otras ubican a estos organismos en una condición ancestral dentro de los decápodos. Siguiendo con otros grupos, observamos dos mecanismos adaptativos, por un lado ocurre una reducción progresiva del tamaño y estructuras esclerosadas (Carideos); mientras que el otro camino es el aumento del tamaño y en el número de estructuras esclerosadas (Parastácidos, Aéglidos y Braquiuros). Dentro de los decápodos, los cangrejos son los que tiene un mayor desarrollo en tamaño y arquitectura de estos órganos. Al igual que los apéndices involucrados en la alimentación, las estructuras internas están adaptadas y modificadas de acuerdo al tipo de dieta, y al comportamiento implicado en su obtención. Por ejemplo muchas especies que realizan macrofagia tienen una alta quitinización de los osículos, a diferencia de aquellas que filtran el alimento. Así, la morfometría del sistema digestivo de los decápodos depende de su historia evolutiva, pero puede ser modificada por varios factores. Además, en estos crustáceos existen varias maneras de adaptarse a las condiciones bióticas y abióticas del ambiente. Por ejemplo hay camarones que sin modificar su anatomía pueden realizar el proceso alimentario de manera eficiente ingiriendo junto con el alimento pequeños granos de arena o roca que son utilizados para macerar las partículas (Collins & Paggi, 1998; Collins, 1999; Williner y Collins, 2002 b)
La digestión comienza con el aporte de enzimas del hepatopáncreas a las cámaras del estómago en donde se realiza la maceración, culminando el proceso digestivo en la glándula del intestino medio (Collins, 2000 b). Una característica a tener en cuenta al hablar de alimentación es el concepto de valores nutricionales y la energía que aportan las presas ingeridas. La característica nutricional de una presa no está solamente dada por sus componentes químicos sino también por otros tópicos. Así, el valor nutricional incluye la facilidad de captura, característica relacionada con el tamaño y la forma de la presa, la digestibilidad, el contenido energético y el contenido de nutrientes esenciales que son característicos para cada organismo depredado.
Las estrategias alimentarias están influenciadas por el pasado evolutivo de las especies. A pesar de este grado de determinismo las taxas poseen un cierto margen de flexibilidad. Todos los animales encuentran en su entorno una gran variedad de potenciales presas, dependiendo de variaciones diarias, estacionales o anuales. Así, dentro de los límites que su pasado evolutivo le impone, un animal puede “seleccionar” una presa u otra, y optar en la utilización de su tiempo y espacio ecológico. Estas “decisiones” tienen sus consecuencias, así las presas más nutritivas, son parte de la dieta de varios consumidores, y la utilización de un mismo recurso por varios predadores provocan la escasez del mismo. Por otro lado, existen recursos con un bajo nivel nutritivo, no siendo buscado por muchos predadores y por lo tanto son abundantes y disponibles. En este caso si el alimento es de baja calidad el animal deberá invertir más tiempo en contacto con el alimento realizando su búsqueda. En los sistemas de agua dulce, como los ambientes del valle aluvial del río Paraná, existen variaciones diarias de los valores nutricionales del alimento, por ejemplo para aquellas especies que ingieren algas debido a los efectos de fotosíntesis y respiración. Si se compara el contenido estomacal de los organismos y las posibles presas en el ambiente, se pueden observar diferencias y esto se debe a que los predadores son selectivos.
Un rasgo controvertido a tener en cuenta y que aún no se ha resuelto es el tema de la digestibilidad del material ingerido. En general el problema se plantea con el material vegetal, ya sea de macrófitas o algas, que en algunas especies es muy importante. En los decápodos del sistema del Plata, D. pagei, T. borellianus, A. uruguayana, M. borellii, M. jelsii, M. amazonicum, A. paraguayensis, P. argentinus, la fracción vegetal ocupa un lugar en la dieta, variando su proporción e importancia en las distintas especies. Uno de los componentes del material vegetal que es decisivo para los herbívoros u omnívoros que lo ingieren es la cantidad de nitrógeno que este contiene. El contenido de nitrógeno varía de acuerdo a la edad de la planta, las más jóvenes poseen gran cantidad y las senescentes menos. En este caso, sería importante poder definir sobre que parte de las plantas ocurre la depredación. Un caso a destacar es el de D. pagei en el cual el análisis de la dieta natural mostró un alto valor del Índice de Importancia Relativa (IRI) para el ítem de macrófitas. En este ejemplo, los trozos de macrófitas eran verdes sin aspecto de pertenecer a material detrítico (Williner & Collins, 2002 a).
Al analizar el ritmo diario de alimentación es importante tener en cuenta el tiempo que cada tipo de ítem tarda en ser digerido y evacuado. Puesto que lo que analizamos en muestras tomadas en cierto momento pueden pertenecer a presas ingeridas con varias horas de anterioridad (Giri et al ., 2002). En D. pagei los componentes tróficos presentaron variaciones a lo largo de las 24 hs. En los momentos de menor intensidad lumínica se registró la mayor riqueza animal, siendo este ítem volumétrica y numéricamente bajo. La captura de oligoquetos y amebas podría estar relacionada con la ingestión de partículas de arena y sedimento. Las diferencias entre el día y la noche podrían deberse a la disponibilidad local de recursos y a factores endógenos. Además, se podría incrementar el riesgo a la depredación por organismos con mecanismos de captura visual. La influencia de este factor exógeno ha sido registrada en lagunas tropicales (Williner & Collins, 2002 a).
Un caso particular a analizar es el de A. paraguayensis , un camarón de hábitos diferentes a los anteriormente mencionados. Esta especie es planctónica y se desplaza en grupo, al modo de las eufasiaseas marinas, es decir que no está íntimamente relacionado con los ambientes litorales vegetados (Collins & Williner, en prensa). Este camarón es considerado omnívoro comiendo algas, rotiferos y microcrustáceos principalmente. Sin embargo larvas de quironómidos y oligoquetos también conforman su trama trófica, evidenciando una transferencia energética desde las comunidades bentónicas hacia las planctónicas (Collins & Williner, en prensa).
Otros dos camarones como M . borellii y P . argentinus son también considerados omnívoros por depredar distintos niveles de la red trófica, siendo en este caso los elementos animales de la comunidad litoral-bentónica los más frecuentes e importantes (Collins & Paggi, 1998; Collins, 1999).
Por otra parte el cangrejo D . pagei es omnívoro con un importante aporte de componentes vegetales en su dieta, considerándose como un ramoneador de macrofitas (Williner & Collins, 2002 a).
Un aspecto importante de los datos aportados por el análisis de contenidos estomacales es el de poder establecer relaciones tróficas, enmarcando a las especies dentro de una trama. Estos decápodos son un importante componente de las comunidades acuáticas debido a sus densidades y su papel en la transferencia de energía. Varios autores mencionan a los decápodos como parte del espectro trófico de peces, por ejemplo Pimelodus clarias (amarillo), P. albicans (moncholo), Parapimelodus valenciennesi (bagre porteño) (Bonetto et al ., 1963, Cabrera et al ., 1973, Oliva et al ., 1981). Además, estos crustáceos intervienen en el intercambio de materia y energía entre los sistemas acuáticos y terrestre. Este importante rol es desempeñado actuando como depredadores ocasionales de arañas, hormigas y otros insectos; y como presas de especies de aves: Phimosus infuscatus infuscatus (cuervillo cara pelada), Ixobrychus involucris (mirasol común), Trigrisoma lineatum marmoratum (hocó colorado), Ardea alba egretta (garza blanca), Butorides striatus fuscicollis (garcita azulada), Nycticorax nycticorax hoactli (garza bruja), Mycteria americana (tuyuyu), Podiceps major major (macá grande), Phalacrocorax olivaceus olivaceus (biguá), Anhinga anhimga anhinga (biguá víbora), Aramides ypecaha (ipacaá), Pitangus sulfuratus (benteveo), Jacana jacana (gallito de agua), Gallinula chloropus (pollona negra), Agelaius cyanopus cyanopus (varillero negro) (Beltzer, 1983 a, b Beltzer & Paporello, 1984; Beltzer, 1984; Navas, 1991; Bó & Darrieu, 1993 Lajmanovich & Beltzer, 1993; Navas, 1993, 1995). También, la literatura menciona que cangrejos trichodactílidos forman parte del espectro trófico de algunas especies de mamíferos como por ejemplo Lontra longicaudis (lobito de río), Crysozion brachyurus (aguará guazú), Procyon cancrivorus (osito lavador) (Massoia, 1976; Bianchini & Delupi, 1993).
Las especies de la familia Aeglidae han sido mencionadas como presas de caimanes en Brasil, como también ejemplares de aéglidos han sido hallados en estómagos de peces en Entre Ríos.
Reproducción
La maduración ovárica comienza con el aumento de la temperatura pudiendo ocurrir varias puestas en el año. En general eclosionan de los huevos individuos de características que asemejan a pequeños adultos a excepción de A . paraguayensis y P . argentinus que liberan larvas menos desarrolladas. El tiempo que transcurren dentro del huevo es mayor que sus congéneres marinos, siendo por otra parte menor el número de huevos (Boschi, 1981; Collins, 2000 b).
Crecimiento
Como en todos los crustáceos, el crecimiento ocurre de manera discontinua, debido a la exocutícula rígida que se elimina en cada proceso de muda. Así que el crecimiento se debe estudiar a partir de dos parámetros básicos: el tiempo de intermuda y el incremento por muda. Estos elementos varían según la edad, el sexo la alimentación, la calidad del ambiente o la presencia de sustancias xenobióticas, temperatura, estructura poblacional (Collins & Petriella, 1999, Renzulli & Collins, 2000, Williner y Collins, 2003).
A su vez en algunas especies como por ejemplo M . borellii el crecimiento relativo de varias partes del cuerpo muestran diferentes patrones de desarrollo asociados a factores endógenos y exógenos. Esto define fases indiferenciadas con un crecimiento isométrico correspondientes a juveniles y otras diferenciadas para adultos con distintos tipos de crecimiento alométrico. La transición entre las fases ocurre por medio de mudas críticas indicando el inicio de la maduración de las gonadas (Collins & Petriella, 1999), La diferenciación entre los sexos tienen un significado en la defensa territorial, combates, desplazamiento y cortejo (Collins, 2001).
Comportamiento
En la naturaleza existe una mayor proporción de especies que presentan comúnmente poblaciones con un arreglo espacial del tipo contagioso. Una excepción a esto se observó en el camarón P . argentinus , el cual varió su disposición en el espacio según la existencia o el tipo de refugio. En laboratorio se determinó que la ubicación de un individuo de P . argentinus no afecta la presencia de otro. Sin embargo en ambientes naturales, el arreglo areal de esta especie puede estar determinado por las características del hábitat (presencia o no de vegetación), la presencia de especies competidoras y/o depredadores (Williner & Collins, 2000 a, b).
En la especie M . borellii se observa una disposición espacial contagiosa. No obstante en latitudes mas altas, con climas de marcadas diferencias estacionales existe un cambio en el arreglo espacial según el momento reproductivo (Boschi, 1981).
En relación con la presencia de un morfo dominante, P . argentinus muestra una ausencia de jerarquización en las poblaciones lo que se relaciona con una agregación al azar. Mientras que el camarón M . borellii cambia el ritmo de crecimiento de algunos ejemplares indicando el comportamiento agonístico de las poblaciones (Collins & Williner, 2000 a).
Otro aspecto del comportamiento es el referido a los ritmos biológicos que en los crustáceos de agua dulce está influenciado por factores endógenos y exógenos. En el camarón M . jelskii la actividad diaria varía con la estructura etaria y sexual y en relación a la presencia o ausencia de vegetación acuática, respondiendo quizás a la periodicidad propia de las presas y depredadores potenciales (Williner & Collins, 2002 b).
Los cambios en los parámetros físico – químicos y biológicos en los ambientes imprimen en las comunidades ajustes de nivel ecológico y etológico. Tales adaptaciones les permite permanecer temporalmente a través de las dispersiones entre parches locales mediante un eficiente aprovechamiento de los mismos. Estas condiciones ambientales favorecerían las interacciones agresivas que podrían dar origen a cierta competencia interespecífica.
Este tipo de patrón de actividad en los decápodos puede ser explicado en dos dimensiones diferentes actuando conjuntamente. Una de ellas es la dimensión trófica acordada por una mayor disponibilidad de alimento y la otra es la de riesgo a ser depredados manifestando su actividad según la presencia y accesibilidad a refugios, y la presencia de enemigos naturales.
Estas vías involucran modos de interacciones y respuestas a esos mecanismos compensatorios cubriendo cambios en el comportamiento, los procesos evolutivos y la dinámica poblacional. Como consecuencia, los organismos ajustan sus actividades diarias a esos factores. Además, la evolución conjunta determinaría los modelos de una manera óptima disminuyendo cualquier interacción intraespecífica (Collins 1995; 1997 a; 2000 b; Renzulli & Collins, 2000; Williner & Collins, 2002 a, b).
Ecotoxicología
Diferentes sustancias xenobióticas que son utilizadas masivamente y presentan un potencial riesgo a ingresar en los sistemas acuáticos afectan la supervivencia y el crecimiento de los camarones y cangrejos (Williner & Collins, 2003; Collins, no publicado).
Características biológicas de este grupo permitirían su utilización en la evaluación ecotoxicológica de diferentes elementos que podrían ingresar al ambiente acuático, siendo también un buen indicador de estrés ambiental.
Cultivo
Existe suficiente información sobre la capacidad de producción de una especie de camarón ( M . borellii ) que podría ser cultivada con fines comerciales (Collins & Petriella, 1996; Collins, 1997 b, 1999 b)
Otra información sobre el grupo
Otros antecedentes que deben mencionarse son aquellos trabajos referidos a las características morfológicas y morfométricas de machos, hembras y huevos (Ringuelet, 1949 c; Boschi, 1963), los primeros estadios larvales (Boschi, 1961; Menu-Marque, 1973), el ciclo reproductor, desove y su relación con la temperatura (Goldstein & Lauria de Cidre, 1974; Bond & Buckup, 1982), la oogénesis y histología de la gónada (Schuldt, 1980 a, b; 1981; 1984), la composición química y metabolismo lipídico y energético (Menu-Marque & Morales, 1974; Rodrigues Capitulo & Freyre, 1979; Gonzales Baro & Pollero, 1988; 1993; Gonzales Baro et al. , 1990; Gonzales Baro, 1991; Irazú et al. , 1992), la duración de la intermuda y el comportamiento reproductivo (Setz & Buckup, 1977), la influencia de la temperatura sobre la duración de la intermuda (Bond & Buckup, 1988), la dinámica poblacional y demografía (Rodriguez Capitulo & Freyre, 1989; Spivak, 1997), la infección con parásitos (Schuldt, 1984; Schuldt & Damborenea, 1989), evaluación como un elemento de control de larvas de mosquito (Collins, 1998; Giri & Collins, en prensa b, c). Morfología de cangrejos y de ejemplares de Aéglidos (Ringuelet, 1948 a, b, 1949, 1960, 1961; Lopretto, 1976, 1978 a, b, 1979, 1981 a), análisis de dimorfismo sexual en cangrejos (Lopretto, 1981 b), discusión biogeográfica de los decápodos (Ringuelet, 1961; Lopretto & Morrone, 1994; Morrone & Lopretto, 1994, 1995; Morone, 1999, 2001), síntesis de la fauna de decápodos de Argentina (Loretto, 1995)
Bibliografía
BeBeltzer, A.H. 1983 a. Alimentación de la “garcita azulada” ( Butorides striatus ) en el valle aluvial del río Paraná Medio (Ciconiiformes: Ardeidae). Revista Hydrobiologia Tropical ., 16 (2): 203-206.
Beltzer, A.H. 1983 b. Alimentación del “benteveo” ( Phitangus sulphuratus ) en el valle aluvial del río Paraná Medio (Passeriformes: Tyrannidae). Revista de la Asociación de Ciencias Naturales del Litoral 14 (1): 47-52.
Beltzer, A.H. 1984. Ecología alimentaria de Aramides ypecaha (Aves: Rallidae) en el valle aluvial del río Paraná Medio (Argentina). Revista de la Asociación de Ciencias Naturales del Litoral 16 (1): 73-83.
Beltzer, A.H. y Paporello, G. 1984. Alimentación de aves en el valle aluvial del río Paraná. IV Agelaius cyanopus cyanopus Vieillot, 1819 (Passeriformes: Icteridae). Iheringia, Ser. Zool . 62: 55-60.
Bianchini, J.J. y Delupi, L.H. 1993. Mammalia p. 1-79. En: Ageitos de Castellanos, Z. (dir.) Fauna de Agua Dulce de la República Argentina. Vol. 44 (2) (actualización). PROFADU, Buenos Aires.
Bó, N.A. y Darrieu, C.A. 1993. Aves ciconiformes p. 1-59. En: Ageitos de Castellanos, Z. (dir.) Fauna de Agua Dulce de la República Argentina. Vol. 43 (1B) . PROFADU, Buenos Aires.
Bond, G. y Buckup, L. 1982. O ciclo reproductor de Macrobrachium borellii (Nobili, 1896) e Macrobrachium potiuna (Muller, 1880) (Crustaceca, Decapoda, Palaemonidae) e suas relacoes com a temperature. Revista Brasileira de Biologiay 3): 473-483
Bond, G. y Buckup, L.1988. O ciclo da intermuda em Macrobrachium borellii (Nobili, 1896) (Crustacea, Decapoda, Palaemonidae): a influencia da temperatura e do comprimento do animal. Revista Brasileira de Zoologia 5(1):45-59.
Bond, G. y Buckup, L. 1999. Os crustáceos do Rio Grande do Sul . Porto Alegre. Ed. Universidade/UFRGS
Bonetto, A.A. y Wais, I.R. 1995. Southern South American streams and rivers . In: Cushing CE, Cummins KW, Minshall GW, eds., Ecosystems of the World. 22. River and stream ecosystems . Amsterdam, Elsevier, pp. 257-293.
Bonetto, A. A., Pignalberi, C. y Cordiviola, E. 1963. Ecología alimentaria del “amarillo” y “moncholo”, Pimelodus clarias (Bloch) y Pimelodus albicans (Valenciennes) (Pices, Pimelodidae). Physis 24 (67): 87-94.
Boschi, E.E. 1961. Sobre el primer estadio de dos especies de camarones de agua dulce. (Crustacea, Palaemonidae). Primer Congreso Sudamericano de Zoología :69-77.
Boschi, E.E. 1963. Observaciones sobre la morfología externa del camarón Macrobrachium borellii (Nobili, 1896) (Crustacea, Palaemonidae). Physis , 24(67):165-179.
Boschi, E. E. 1981. Decapoda Natantia. Fauna de Agua Dulce de la República Argentina , PROFADU, Buenos Aires, 26: 1-61.
Brusca, R.C. y Brusca, G.J. 1990. Invertebrates . Sinauer Associates, Sunderland Massachusets, 923p.
Cabrera, D.E., Baiz, M.L., Candia, R. y Christiansen, H.E. 1973. Algunos aspectos biológicos de las especies de ictiofauna de la zona de Punta Lara (río de la Plata). 2ª parte. Alimentación natural del bagre porteño ( Parapimelodus valenciennesi ). Armada Argentina, Servicio de Hidrografía Naval, H.1029 , 7-47.
Collins, P. 1995. Variaciones diarias de la actividad trófica en una población de Palaemonetes argentinus (Crustacea Decapoda). Revista de la Asociación de Ciencias Naturales del Litoral 26(1): 57-66.
Collins, P. 1997 a. Ritmo diario de alimentación en el camarón Macrobrachium borellii (Decapoda, Palaemonidae). Iheringia Série Zoologia 82:19—24.
Collins, P.1997 b. Cultivo del camarón Macrobrachium borellii (Crustacea: Decapoda: Palaemonidae), con dietas artificiales. Natura Neotropicalis 28(1): 39-45
Collins, P.A.1998. Laboratory evaluation of the freshwater prawn, Macrobrachium borellii as a predator of mosquito larvae. Aquatic Science 60:22-27.
Collins, P.A. 1999 a. Feeding of Palaemonetes argentinus (Nobili) (Decapoda: Palaemonidae) in flood valley of river Parana Argentina. Journal of Crustacean Biology 19(3): 485-492.
Collins, P.A. 1999 b. Role of natural productivity and artificial feed in enclosures with the freshwater prawn, Macrobrachium borellii (Nobili, 1896). Journal of Aquaculture Tropical 14(1):47-56.
Collins, P.A. 2000 a. A new distribution for Macrobrachium jelskii (Miers, 1877) in Argentina. Crustaceana 73(9): 1167-1169.
Collins, P.A. 2000 b. Mecanismos de coexistencia de poblaciones de palaemónidos dulciacuícolas (Crustacea, Decapoda, Caridea) . Tesis Doctoral Univ. Nac. De La Plata.
Collins, P.A. 2001. Relative growth of the freshwater prawn Macrobrachium borellii (Nobili, 1896) (Decapoda: Palaemnidae) . Nauplius 9(1): 53-60.
Collins, P.A. y Paggi, J.C. 1998. Feeding ecology of Macrobrachium borellii (Nobili) (Decapoda: Palaemonidae) in flood valley of river Parana Argentina. Hidrobiologia 362:21-30.
Collins, P.A. y Petriella, A 1996. Crecimiento y supervivencia del camarón Macrobrachium borellii (Decapoda: Palaemonidae) alimentado con dietas artificiales. Neotropica 42(107y108): 3-8.
Collins, P.A. y Petriella, A. 1999. Growth Pattern of isolated prawns of Macrobrachium borellii (Crustacea, Decapoda, Palaemonidae). Inertebrate. Reproduction and Development 36:1-3.
Collins, P.A. y Williner, V. 2003. Feeding of Acetes paraguayensis (Nobili) (Decapoda: Sergestidae) in flood valley of river Paraná Argentina. Hydrobiologia 493: 1-6.
Collins, P.A., Williner, V. y GIRI, F. 2002. A new distribution record for Zilchiopsis oronensis (Miers, 1877) in Argentina. Crustaceana 75(7): 931-934.
Fernandez, D. y Collins, P. en prensa. Estrategia de supervivencia de cangrejos en ambientes dulciacuícolas inestables. Natura Neotropicalis .
Giri, F. y Collins, P. 2003. Evaluación de Palaemonetes argentinus (Decapoda: Natantia) (Nobili, 1901) como controlador biológico de larvas de mosquito Culex pipiens s.l. (Diptera: Culicidae) en condiciones de laboratorio. Iheringia Serie Zoologica 13(3): 237-242.
Giri, F. y Collins, P. en prensa a. A geometric morphometric analysis of two sympatric species of family Aeglidae (Crustacea, Decapoda, Anomura) from La Plata basin. Journal of Italian Zoology
Giri, F. y Collins, P. en prensa b. Eficiencia de captura del camarón dulciacuícola Palaemonetes argentinus (Nobili 1901) sobre larvas de mosquito Culex pipiens s.l. en laboratorio. Hidrobiologia .
Giri, F., Williner, V. y Colllins, P. 2002. Tiempo de evacuación del camarón dulceacuícola Palaemonetes argentinus (Crustacea, Decapoda) alimentado con larvas de mosquito Culex pipiens s.l.. FABICIB 6: 37-41.
Goldstein, B. y Lauria de Cidre, L. 1974. Ciclo de maduración sexual y observaciones preliminares sobre el desove del camarón dulceacuicola Palaemonetes argentinus (Nobili, 1901) (Crustacea, Decapoda, Palaemonidae) I Hembra. Physis B 33(87): 165-176.
Gonzalez Baro, M. del R. 1991. Composición y metabolismo del camarón Macrobrachium borellii . Tesis U.N.La Plata. Fac. de Cs. Nat. y Museo.
Gonzalez-Baro, M. del R., Irazu, C. y Pollero, R. 1990. Palmitoyl-CoA ligase activity in hepatopancreas and gill microsomes of the freshwater shrimp Macrobrachium borellii . Comparative Biochemistry and Physiology. B 97:129-133
Gonzalez-Baro, M.del R. y Pollero, R. 1988. Lipid characterization and distribution among tissues of the freshwater crustacean Macrobrachium borellii during an annual cycle. Comparative Biochemistry and Physiology B 98:129-133.
Gonzalez-Baro, M. del R. y Pollero, R. 1993. Palmitic acid metabolism in hepatopancreas of the freshwater shrimp Macrobrachium borellii . Comparative Biochemistry and Physiology. B 106(1):71-75.
Irazu, C.E., Gonzalez Baro, M. del R. y Pollero, R. 1992. Effect of environmental temperature on mitochondrial B-oxidation activity in gills and hepatopancreas of the freshwater shrimp Macrobrachium borellii . Comparative Biochemistry and Physiology 102B(4):721-725.
Lajmanovich, R. C. y Beltzer, A.H. 1993. Aporte al conocimiento de la biología alimentaria de la pollona negra Gallinula chloropus en el Paraná Medio, Argentina. El Hornero 13 (4): 289-291.
Lopretto, E.C. 1976. Morfología comparada de los pleópodos sexuales masculinos en los Trichodactylidae de la Argentina (Decapoda, Brachyura). Limnobios 1(3): 67-94.
Lopretto, E.C. 1978a. Estructura exoesqueletaria y miológica del quinto par de periópodos del macho de la familia Aeglidae (Crustacea Anomura). Limnobios 1(8): 284-298.
Lopretto, E.C. 1978b. Las especies de Aegla Leach del centro-oeste argentino en base a la morfología comparada del quinto par de periópodos (Crustacea, Anomura, Aeglidae). Neotropica 24(71): 57-68.
Lopretto, E.C. 1979. Estudio comparativo del quinto par de pereiópodos en los representantes del género Aegla de la Patagonia argentina (Crustacea, Anomura). Neotrópica 25(73): 9-22.
Lopretto, E.C. 1980. Análisis de las características del quinto pereiópodo en las especies de Aegla del grupo “ platensis ” (Crustacea, Anomura, Aeglidae). Physis B 39(96): 37-56.
Lopretto, E.C. 1981a. Consideraciones sobre la estructura apendicular vinculada al dimorfismo sexual en los machos de las especies de Aegla del noroeste argentino. (Crustacea, Anomura, Aeglidae). Acta zoológica Lilloana 36(2): 15-35.
Lopretto, E.C. 1981b. Discusión sobre las presuntas subespecies de Dilocarcinus (D.) pagei (Crustacea Brachyura Trichodactylidae). Redescripción y referencia a su polifenismo. Physis B 39(97): 21-31.
Lopretto, E.C. y Morrone, J.J. 1994. Areas de endemismo de decápodos dulceacuícolas (Crustacea: Malacostraca) de América del Sur Austral. I Congreso y III Reunión Argentina de Limnología. Tankay 1: 100-102.
Lopretto, E.C. 1995. Crustacea Eumalacostraca , pp. 1001-1039. En: Lopretto, E. C. & G. Tell (dir.). Ecosistemas de Aguas Continentales. Tomo III. Ediciones Sur.
Magalhães, C. y Türkay, M. 1996 a. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica 75(1/2): 63-95.
Magalhães, C. y Türkay, M.1996 b. Taxonomy of the Neotropical freshwater crab family Trichodactylidae II. The genera Forsteria, Melocarcinus, Sylviocarcionus , and Zilchiopsis (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica 75(1/2): 97-130.
Magalhães, C. y Türkay, M. 1996 c. Taxonomy of the Neotropical freshwater crab family Trichodactylidae III. The genera Fredilocarcinus and Goyazana (Crustacea: Decapoda: Brachyura. Senckenbergiana biologica 75(1/2): 131-142.
Manning, R.B. y Hobbs, H.H. 1977. Decapoda . In: Hurlbert H, ed., Biota acuática de Sudamérica Austral , San Diego, California, San Diego State University, pp. xiv + 342.
Martin, J.W. y Davis, G.E. 2001. An update classification of the recent Crustacea. Science Series (Natural History Museum of Los Angeles County) Los Ángeles, 39: 1-124.
Massoia, E. 1976. Mammalia p. 1-128. En: Ringuelet, R. A. (dir.) Fauna de Agua Dulce de la República Argentina. Vol. 44 . FECIC, Buenos Aires.
Menu-Marque, S. 1973. Desarrollo larval de Palaemonetes argentinus (Nobili, 1901) en el laboratorio (Crustace, Caridea, Palaemonidae). Physis B 32(85): 149-169.
Menu-Marque, S. y Morales, J.E. 1974. La composición química del camarón Macrobrachium borellii (Nobili 1896) (Crustacea, Caridea, Palaemonidae). Physis B 33(87): 195-199.
Morrone, J.J. 1999. Presentación preliminar de un nuevo esquema biogeográfico de América del Sur. Biogeographica 75 (1): 1-16.
Morrone, J.J. 2001. A proposal concerning formal definitios of the neotropical and andean regions. Biogeographica 77 (2): 65-82.
Morrone J.J. y Lopretto, E.C. 1994. Distributional patterns of freshwater Decapoda (Crustacea: Malacostraca) in southern South America: a panbiogeographic approach. Journal of Biogeography 21: 97-109.
Morrone J.J. y Lopretto, E.C. 1995. Parsimony analysis of endemicity of freshwater Decapoda (Crustacea: Malacostraca) from southern South America. Neotropica 41 (105-106): 3-8.
Morrone J.J. y Lopretto, E.C. 2001. Trichodactylid biogeographic patterns (Crustacea: Decapoda) and the Neotropical region. Neotrópica 47: 49-55.
Navas, J. 1991. Aves gruiformes p. 1-80. En: Ageitos de Castellanos, Z. (dir.) Fauna de Agua Dulce de la República Argentina. Vol. 43 (3) . PROFADU, Buenos Aires.
Navas, J. 1993. Aves Podicipediformes y Pelecaniformes p. 1-79. En: Ageitos de Castellanos, Z. (dir.) Fauna de Agua Dulce de la República Argentina. Vol. 43 (1A) . PROFADU, Buenos Aires.
Navas, J. 1995. Aves ciconiformes p. 1-53. En: Ageitos de Castellanos, Z. (dir.) Fauna de Agua Dulce de la República Argentina. Vol. 43 (1C) . PROFADU, Buenos Aires.
Oliva, A., Ubeda, C.A., Vignes. E.I. y Iriondo, A. 1981. Contribución al conocimiento de la ecología alimentaria del bagre amarillo ( Pimelodus maculatus Lacépède 1803) del río de la Plata (Pisces, Pimelodidae). Comunicaciones del Museo Argentino de Ciencias Naturales Ecología 1 (4): 31-50.
Pettovello, A.D. 1996. First record of Macrobrachium amazonicum (Decapoda, Palaemonidae) in Argentina. Crustaceana 69(1):113-114.
Renzulli, P. y Collins P. 2000. Influencia de la temperatura en el crecimiento del cangrejo Trichodactylus borellianus. FABICIB 4: 129-136.
Renzulli, P. y Collins P. 2001. Ritmo nictimeral de la actividad locomotora de los cangrejos dulciacuícolas Dilocarcinus pagei pagei y Trichodactylus borellianus. FABICIB 5:145-153.
Ringuelet, R.A. 1948a. Una nueva Aegla del Nordeste Argentino (Decapoda, Anomura). Notas Museo La Plata 13: 203-208.
Ringuelet, R.A. 1948b. Los “cangrejos” argentinos del género Aegla de Cuyo y la Patagonia. Revista del Museo La Plata (N.S.), Zoología 5: 297-347.
Ringuelet, R.A. 1949 a. Camarones y cangrejos de la zona de Goya (Sergéstidos, Palamonidae y Trichodactylidae). Notas del Museo de La Plata Zoología 14 (119): 79-109.
Ringuelet, R.A. 1949 b. Los anomuros del género Aegla del noroeste de la República Argentina. Revista del Museo de La Plata (N.S.), Zoología . 6: 1-45.
Ringuelet, R.A. 1949 c. Consideraciones sobre las relaciones filogenéticas entre las especies del género Aegla Leach (Decapodos Anomuros). Notas Museo La Plata, Zoología 14: 111-118.
Ringuelet, R.A. 1949 d. La prioridad en el estudio biométrico de los Decapodos Anomuros del género Aegla Leach. Notas Museo La Plata, Zoología 14: 119-121.
Ringuelet, R.A. 1960. Identificación de los crustáceos anomuros del género Aegla de la República de Bolivia. 1º Congreso Sudamericano de Zoología, La Plata , Actas y Trabajos , secc. 3 2: 245-249.
Ringuelet, R.A. 1961 a. Notas sobre Aegla de Argentina y Paraguay (Crust. Decap. Anomura). Physis 21 (61): 231-239.
Ringuelet R.A. 1961 b. Rasgos fundamentales de la zoogeografía de la Argentina. Physis 22(63):151-170.
Rodríguez, G. 1981. Decapoda . In: S.H. Hurlbert, G. Rodríguez & N.D. Santos (eds.), San Diego State University, San Diego, California, Aquatic biota of tropical South America, I: Arthropoda, i – xii, 1-323.
Rodríguez, G. 1992. The freshwater crabs of America. Family Trichodactylidae and supplement to the family Pseudothelphusidae. Faune Tropicale 31: 1-189.
Rodrigues Capitulo, A. y Freyre, L. 1979. Metabolismo energético del camarón de agua dulce Palaemonetes argentinus Nobili (Decapoda Natantia Caridea Palaemonidae) de la laguna Chascomús. Limnobios 1(9): 337-345.
Rodrigues Capitulo, A. y Freyre, L. 1989. Demografia de Palaemonetes ( Palaemonetes ) argentinus Nobili (Decapoda Natantia). I Crecimeinto. Limnobios 2(10): 744-756.
Schmitt, W. L. 1942. The species of Aegla , endemic South American fresh-water crustaceans. Proceeding United States Natural Museum 91: 431-520.
Schuldt, M. 1980 a. Evolución de la célula sexual femenina de Palaemonetes argentinus Nobili, 1901 (Crustacea Palaemonidae) durante la ogénesis. Limnobios 1(10): 461-468
Schuldt, M. 1980 b. Acerca de la presencia en Palaemonetes argentinus Nobili (Crustacea Palaemonidae) de elementos germinales asimilables a las células de Bindford. Physis B 39(96): 23-26.
Schuldt, M. 1981. Interpretación funcional de cambios microanatómicos en el ovario de Palaemonetes argentinus Nobili 1901 (Crustacea Palaemonidae) durante desove y post puesta. Limnobios 2(3): 141-151.
Schuldt, M. 1984. Alteraciones de la función reproductora de Palaemonetes argentinus (Crustacea, Palaemonidae) ocasionadas por la infestación con larvas de un probable Phyllodistomun sp. (Trematoda Digenea). Limnobios 2(8): 646-651.
Schuldt, M. y Damborenea, M.C. 1989. Infeccion de Palaemonetes argentinus (Crustacea Palaemonidae) con Probopyrus cf. oviformis (Crustacea Bopyridae) en el canal Villa Elisa (Selva marginal de Punta Lara, Provincia de Buenos Aires, Argentina) I. Estructura poblacional del consorcio, interaccion y fluctuacion. Biota 5: 21-53.
Setz, E. y Buckup, L. 1977. A Duracao da intermuda e o comportamento reprodutivo de Macrobrachium borellii (Nobili, 1896) e Palaemonetes ( Palaemonetes ) argentinus Nobili, 1901 no cultivo em laboratorio (Crustacea, Decvapoda, Palaemonidae). Revista Brasileira de Biologia 37(4): 899-906.
Spivak, E.D. 1997. Life history of a brackish-water population of Palaemonetes argentinus (Decapoda: Caridea) in Argentina. Annales de Limnologie 33(3): 179-190.
Williner V. y Collins, P. 2000 a. ¿Existe jerarquización en las poblaciones de Palemónidos del valle aluvial del Río Paraná?. Natura Neotropicalis 31(1y2): 53-60.
Williner V. y Collins, P. 2000 b. Observaciones en laboratorio de la actividad diaria del cangrejo Dilocarcinus pagei pagei (Brachyura: Tricodactylidae). VII Jornadas de Ciencias Naturales del Litoral. Santa Fe p: 173.
Williner V. y Collins, P. 2002 a. Daily rhythm of feeding activity of a freshwater crab Dilocarcinus pagei pagei in National Park Río Pilcomayo, Formosa, Argentina . In: Modern approaches to the study of Crustacea . Escobar –Briones & Alvarez (eds). Kluver Academic and Plenum Publishers.
Williner V. y Collins, P. 2002 b. Variación espacio-temporal de la actividad del camarón Macrobrachium jelskii (Miers, 1877). Ecología Austral 12:3-10.
Williner V. y Collins, P. 2003. Effects of cypermethrin upon the freshwater crab Trichodactylus borellianus (Crustacea: Decapoda: Braquiura). Bulletin of Environmental Contamination and Toxicology 71(1): 106-113.
Tabla 1. Davis (2001). Algunas subdivisiones en el Infraorden Brachyura fueron suprimidas para una más fácil compilación de los registros.
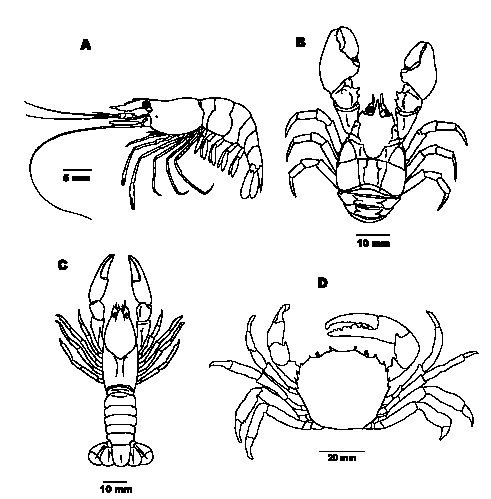
Fig. 1. A) Macrobrachium borellii; B)Begla platensis; C)Parastacus pilimanus; D) Zilchiopsis collastinensis (modificado de Buckup & Bond-Buckup, 1999)