Biodiversidad de Porifera en el Litoral Argentino. Grado de Competencia con el Bivalvo invasor Limnoperna fortunei (Dunker, 1857) (Bivalvia, Mytilidae)
Inés EZCURRA de DRAGO1
1Instituto Nacional de Limnología - INALI - (CONICET-UNL). J. Macia 1933, 3016 Santo Tomé, Prov. Santa Fe, Argentina. E-mail: inesezcurra@arnet.com.ar
Abstract : Biodiversity of Porifera of the Argentina Mesopotamia. The freshwater of the world are included in seven families. Three of them are represented in the Mesopotamian Region of Argentina (where portions of the catchment areas of the Paraná, Uruguay and Río de La Plata river are developed): Spongillidae Gray, Metaniidae Volmer-Ribeiro and Potamolepidae Brien. In aggreement with others South American freshwater invertebrates taxa, the sponges show a considerable number of endemic species and genera. Spongillidae, the only cosmopolitan family, is represented only by three species and Potamolepidae, restricted to ancient large hidrographic basins of the Afrotropical and Neotropical Regions, has a hicgh value of species richness.
The geographic distribution of Argentine freshwater sponges shows a remarkable heterogeneity, being the specific richness and the highest density in the Mesopotamian region. The recent invasion of Limnoperna fortunei (Bivalvia, Mytilidae) in the Mesopotamian rivers determine a spatial competence of this exotic species with the sponges, since L. fortunei has been founded in the same sustrate that them. This fact could determine the lowering diversity of the sponges developed in the above mentioned fluvial hydrosystems.
Key words: Freshwater porifera, Mesopotamia argentina, Geographic distribution.
Palabras clave: Porisfera agua dulce, Mesopotamia argentina, distribución geográfica.
Introducción
Todas las esponjas de los ambientes epicontinentales pertenecen al Suborden Spongillina Manconi y Pronzato, 2002 (Demospongiae, Haplosclerida). Debido a la composición silícea de su sistema esquelético de gran perdurabilidad, las esponjas dulciacuícolas constituyen un Phylum de especial interés no solamente para estudios de neodiversidad sino también de paleodiversidad. No obstante, la mayor parte de las especies dulciacuícolas tienen registros fósiles no muy antiguos, existiendo una especie Paleártica, ( Spongilla gutenbergiana Muller et al . (1982) y dos especies Sudamericanas ( Palaeospongilla chubutensis Ott y Volkheimer, 1972 y Spongilla patagonica Volkmer-Ribeiro y Reitner, 1991) del Cretáceo Inferior.
Si bien algunos autores han señalado gran adaptabilidad de estos organismos a distintos tipos de ambientes epicontinentales con diversos gradientes tróficos, debe destacarse que solamente un escaso número de especies son eurioicas, presentando la mayor parte de ellas una marcada fidelidad ecológica demostrada en el estudio de las especies de la fauna actual así como en aquellas registradas en estudios estratigráficos (Racek 1966, 1970, 1974; R(Racek & Harrison 1974; Harrison & Warner 1986; Harrison 1988). Además, tanto en estudios experimentales como en investigaciones desarrolladas en ambientes de características limnológicas físicas y químicas muy diversas, se han determinado variaciones morfométricas de gemoscleras y macroscleras e, incluso, malformaciones de ambos tipos de espículas como respuesta a determinados parámetros (Jewell 1935, Poirrier 1969, 1974; Harrison 1974, Moore 1953, Ezcurra de Drago 1972, 1975, 1976, 1988, 1995). Por tal motivo, se las considera especies indicadoras de calidad del agua (Ezcurra de Drago 1995).
Al presente son reconocidas 190 especies, agrupadas en siete familias (Manconi y Pronzato 2002): Spongillidae Gray, Lubomirskiidae Rezvoj, Malawispongiidae Manconi y Pronzato, Metaniidae Volkmer-Ribeiro, Metschnikowiidae Czerniavsky, Potamolepidae Brien y Palaeospongillidae Volkmer Ribeiro y Reitner, siendo esta última fósil.
El Phylum presenta en general un elevado grado de endemismo. Además, y como ha sido observado para otros grupos zoológicos (Ringuelet 1961; Rapoport 1968; Fittkau 1969), en las esponjas de la región Neotropical se registra, asimismo, un elevado número de taxa endémicos tanto a nivel de géneros como de especies. Del total registrado, un 74 % de las especies son endémicas, lo que señala la evolución del grupo en esta región, considerada por los citados autores como centro de origen y dispersión de numerosos taxa.
Tanto la superficie (19.200.000 km 2 ; Rapoport, 1968), como la gran extensión latitudinal (12º N - 50 º S) de la región Neotropical, permiten la existencia de un gradiente climático (desde tropical a frío), variaciones hipsométricas, hidrográficas, fisiográficas y ecológicas, las que determinan una gran heterogeneidad de ambientes, posibilitando una mayor diversidad específica (Blondel 1986, Ezcurra de Drago, 1993). A ello debe sumarse, para la región Mesopotámica, la presencia de grandes ríos con extensas llanuras aluviales como los tramos medio e inferior del Paraná, los que brindan a las esponjas una amplia diversidad de hábitats a distintos niveles verticales en la vegetación de la zona de transición acuático-terrestre (ATTZ, Junk et al. 1989).
Debe señalarse que, a excepción de algunos géneros monoespecíficos y/o especies de distribución muy reducida, los grandes sistemas fluviales Sudamericanos (Amazonas, Orinoco, Magdalena, Paraguay, Paraná y Uruguay), presentan un elevado número de especies en común. Ello evidencia el rol de los citados ríos como corredores ecológicos. En efecto, y tal como expresan Soldano (1947), Morello (1984) y Vokmer Ribeiro (2000), durante las grandes crecientes, los cursos de las cabeceras del Orinoco se comunican con aquellos similares del Amazonas, los que a su vez se conectan en los períodos de crecientes con las nacientes del Paraguay Superior. Ello explica la alta similitud existente entre la fauna de Poríferos de los grandes sistemas hidrográficos de Sudamérica (Ezcurra de Drago, 1993, Volkmer Ribeiro 2000, Manconi y Pronzato 2002).
El tipo de reproduccion sexual y/o asexual desarrollado por las esponjas en las aguas epicontinentales durante el proceso evolutivo, explican parte de la causalidad del alto grado de endemismo de algunas familias y/o géneros y/o especies y los muy escasos taxa de distribución cosmopolita. En efecto, el desarrollo de cuerpos de resistencia (gémulas) con una estructura compleja altamente especializada para lograr una dispersión pasiva observado en las especies de Spongillidae y Metaniidae, les ha permitido una amplia distribución geográfica. En Potamolepidae en cambio, las gémulas son escasas, de desarrollo incrustante sobre el sustrato y por ende, con pocas posibilidades de dispersión. Lubomirskidae, Malawispongiidae y Metschnikowiidae, típicas de grandes lagos, con un tipo de vida «perenne» y reproducción exclusivamente sexual, pueden extender su distribución solamente a través de las larvas; siendo estas las familias que presentan el mayor grado de endemismo.
En cuanto a su distribución, las especies de Porifera registradas hasta el presente en Argentina (Ezcurra de Drago, 1993) ocupan dos grandes áreas bien diferenciadas: 1) el sistema de los grandes ríos de la Cuenca del Río de la Plata, Paraná (incluyendo los ambientes de su extensa llanura aluvial), Paraguay, Uruguay y algunos tributarios), y el estuario del Río de la Plata es decir un área geógrafica comprendida dentro de la subregión Chaqueña, provincias de Chaco y Pampa (Morrone, 2002) (o dominio Subtropical de la subregión Guayano-Brasileña (Ringuelet, 1961), y 2) las aguas comprendidas en el amplio territorio existente en el centro, oeste y sur del país, que corresponde a las provincias de Patagonia Central y Patagonia Subandina de la subregión Patagónica (Morrone, 2002) (o al dominio Andino-Patagónico y Pampásico de la Subregión Guayano-Brasileña; Ringuelet, 1961). La definición del límite entre las citadas regiones zoogeográficas, varía en relación con los autores y con los taxa de referencia (Rapoport 1968). Para el caso que nos ocupa, el límite sur de la región Guayano-Brasileña está dado por el estuario del Río de la Plata.
Debe señalarse que si bien han sido mucho más numerosos los muestreos efectuados en los ambientes de la Cuenca del Río de la Plata, se han realizado también muestreos extensivos en el resto del país, incluyendo ambientes lacustres de la Antártida, lo que permite afirmar que las diferencias mencionadas respecto a la heterogeneidad espacial de la riqueza de especies del Phylum en nuestro país están basadas en información fidedigna.
Estado actual del conocimiento de las esponjas de la Mesopotamia
Si bien el conocimiento de la sistemática y distribución de las especies de ambientes epicontinentales argentinos está lejos de ser exhaustivo, proporciona una información bastante amplia.
Treinta y tres son las especies de esponjas actuales registradas para Argentina, representando un 64 % del total de especies halladas hasta el presente en la región Neotropical. Están comprendidas en tres de las familias de mayor distribución a nivel mundial: Spongillidae, Potamolepidae y Metaniidae. Una cuarta familia es Palaespongillidae Volkmer-Ribeiro y Reitner, monoespecífica y fósil, presente solamente en la Patagonia.
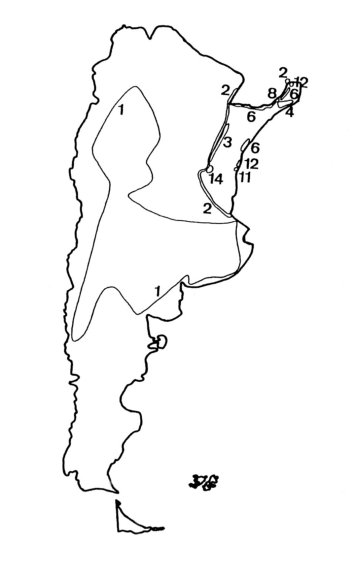
Spongillidae Gray, es la única familia cosmopolita, existiendo en la región litoral uno de los pocos taxa cosmopolitas a nivel de especie: Eunapius fragilis Leidy. De los 21 géneros válidos a nivel mundial, 9 están representados en Argentina y 8 en la región mesopotámica. Es la familia que posee el mayor número de especies, especialmente en ambientes del área de transición acuático terrestre (ATTZ), de la llanura aluvial del Paraná Medio (Fig. 1, Ezcurra de Drago, 1993), con mayor grado de variación de la morfología externa de los ejemplares, así como de la estructura de sus microscleras y gemoscleras. Las gémulas poseen, en general, una gruesa capa neumática que facilita su flotación y por ende, su dispersión y colonización de nuevos sustratos. La mayor parte de las especies son perifíticas y/o pleustónicas, existiendo solamente algunas de hábitos bentónicos.
Metaniidae Volkmer-Ribeiro, se encuentra en las regiones Neártica, Neotropical, Afrotropical, Oriental y Australiana. De los cinco géneros que posee la familia a nivel mundial e incluso en la región Neotropical, en la Mesopotamia se encuentran representados solamente dos de ellos. El de mayor distribución es Drulia Gray, representado por dos especies de muy distintos requerimientos ecológicos (Tabla I) y Houssayella Bonetto y Ezcurra de Drago, monoespecífico, de hábitat bentónico y restringido a ambientes fluviales ritrónicos. Tanto las microscleras como las gemoscleras de las especies de Metaniidae son, al igual que las de Spongillidae, de morfología muy variable, siendo las del género Drulia las que se diferencian en mayor grado de las restantes especies de ambas familias, por la estructura en forma de escudos de sus gemoscleras.
Potamolepidae Brien, se encuentra distribuida en las antiguas y extensas cuencas hidrográficas de las regiones Afrotropical (ríos Congo, Zambesi, etc.) y Neotropical (los grandes ríos Sudamericanos). De los 6 géneros válidos de la familia, en la región Mesopotámica se encuentran representados dos, Uruguaya Hinde y Oncosclera Volkmer-Ribeiro, .
A diferencia de las dos familias anteriores, Potamolepidae posee solamente una especie, Oncosclera navicella, la que si bien es predominantemente bentónica, suele registrársela en el perifiton en la ATTZ de la llanura aluvial del Paraná Medio durante las fases de inundación. El resto de las especies son estrictamente bentónicas, generalmente epilíticas, desarrolladas en ríos de cabecera tipo ritron. La estructura gemular, típica de la familia, se caracteriza por poseer gémulas de tipo adherentes al sustrato, desprovistas en la mayor parte de las especies de la capa neumática presente en las dos familias anteriores y de tubo foraminal. No obstante, estudios efectuados sobre las variaciones ecomórficas de Oncosclera navicella han demostrado que la estructura de las gémulas de esta especie presentan marcadas variaciones en relación a las características hidrológicas del hábitat (Ezcurra de Drago, inédito).
Los resultados obtenidos hasta el presente, señalan que, si bien los valores de riqueza específica son casi idénticos, existe una baja similitud cualitativa entre las especies desarrolladas en el Paraná Superior, algunos de sus tributarios y los tramos Superior y Medio del río Uruguay, respecto a las registradas en la llanura aluvial del río Paraná Medio. Las diferencias cualitativas señaladas, se explican por la fidelidad ecológica que poseen gran parte de las especies de Porifera señaladas anteriormente. En efecto, podríamos referirnos a especies que se adaptan a tramos ritrónicos y otras a sectores del potamon. En el primer caso, las especies son bentónicas, de desarrollo incrustante epilítico, mesohilo de trama cerrada, con ósculos a veces imperceptibles a simple vista. Están adaptadas a aguas rápidas, claras, con gémulas adherentes al sustrato, las que son desarrolladas ya en organismos muy pequeños (de hasta 3 mm de diámetro), adaptándose de esta forma a ríos de régimen torrencial, como es el caso de los tributarios del Paraná Superior, en la provincia de Misiones, incluyendo los distintos saltos de las Cataratas del Iguazú. En estos hábitats son dominantes, tanto por la riqueza específica como por su biomasa, las especies de Potamolepidae. En el segundo caso, los altos valores de riqueza específica registrados están dados fundamentalmente por especies que colonizan, durante las inundaciones, la vegetación palustre y terrestre de la zona de transición acuático-terrestre de la llanura aluvial del Paraná Medio(ATTZ, Junk et al., 1989). Durante crecientes del río Paraná de gran magnitud y larga duración, se ha detectado una densidad elevada de individuos pertenecientes casi en su totalidad a las familias Spongillidae y Metaniidae. En efecto, la única especie de Potamolepidae que suele desarrollarse en los citados sustratos es Oncosclera navicella , como se ha dicho anteriormente. Las especies desarrolladas sobre la vegetación de la ATTZ, poseen todas el mesohilo con trama abierta, desarrollo exuberante, ósculos prominentes, gémulas siempre muy numerosas, dispuestas entre los haces de macroscleras, con gruesa capa neumática la que les permite flotar y colonizar de esta forma otros sustratos.
Competencia entre las especies de Porifera y Limnoperna fortunei
Los estudios realizados hasta el presente sobre la diversidad y distribución de las esponjas, se llevaron a cabo sobre materiales colectados antes del arribo al Paraná Medio del bivalvo invasor Limnoperna fortunei (Dunker, 1857) (Bivalvia, Mytilidae). Esta especie, detectada por primera vez en el estuario del Río de la Plata (Pastorino et al. 1993), debido a sus estrategias adaptativas tuvo una rápida dispersión aguas arriba (Darrigran y Ezcurra de Drago 2000), habiéndose constatado durante los grandes períodos de creciente, especialmente en el Niño 1997-98, un exuberante desarrollo en la vegetación palustre y terrestre de la llanura aluvial del río Paraná Medio. En tal período se registraron poblaciones de L. fortunei bisadas entre el mesohilo de una de especie de Trochospongilla Vejdovsky en un tronco de Solanum glaucophyllum (Ezcurra de Drago, inédito).
L. fortunei , de abolengo marino, posee fecundación externa, con larva tipo veliger, planctónica, la que en su último estadio (plantígrada), ya posee biso, estructura que conserva durante toda su vida, siendo por ende epifaunal. Ello marca una gran diferencia con respecto al tipo de desarrollo larval de las especies autóctonas, las que en su mayor parte son endobentónicas.
El resultado de los estudios llevados a cabo sobre L. fortunei , que incluyen observaciones de campo así como experiencias de labotario, señalan que esta especie, bisada desde el momento de su implantación en el estadio de plantígrada, ocupa idénticos tipos de sustratos colonizados por las esponjas. Ello es válido tanto para las especies de Porifera epilíticas de los tramos superiores de los ríos Paraná y Uruguay, incluyendo los tributarios, con características de «ritron», como para las perifíticas de las zonas del «potamon».
Hasta el presente solamente se han expuesto, a manera de hipótesis, la disminución de la diversidad de bivalvos autóctonos que podría producir la invasión de L. fortunei . Si consideramos el hábito epifaunal de esta especie, respecto al hábito endobentónico de la mayor parte de los bivalvos autóctonos, la competencia en este caso sería desde el punto de vista trófico. En el caso de las esponjas, podría existir no solamente una competencia por el alimento, sino también por el sustrato.
Investigaciones que se consideran necesario realizar
- Analizar la riqueza específica de esponjas de los ambientes fluviales ya estudiados anteriormente a la invasión de Limnoperna fortunei , con el fin de determinar posibles variaciones en la riqueza específica posteriores al ingreso de dicha especie de bivalvo.
- Analizar la coexistencia de especies de esponjas y Limnoperna fortunei en un mismo sustrato o
sustratos similares.
- Comparar la abundancia de ambos taxa.
- Estudiar la riqueza específica de esponjas en sectores de la llanura aluvial del río Paraná aún no investigados, como el tramo inferior y el delta.
- Estudiar la riqueza específica de esponjas de tributarios de los tramos superiores de los ríos Paraná y Uruguay.
- Profundizar los conocimientos acerca de la fidelidad ecológica de los taxa de esponjas.
Las investigaciones se realizarán, en parte, mediante el análisis de futuros muestreos así como sobre materiales existentes en las colecciones del INALI, especialmente en lo referido a las especies de esponjas. Estos materiales proceden de tributarios del Paraná Superior, en la provincia de Misiones, del río Iguazú, de la represa Yacyretá y del Uruguay Superior. Todos los materiales citados fueron colectados previamente al ingreso de Limnoperna fortunei a la cuenca del Río de la Plata.
El análisis de la posible competencia de L. fortunei sobre la diversidad de esponjas, se llevará a cabo sobre materiales recolectados en fases de estiaje, en períodos inmediatamente posteriores al descenso de las aguas, para evitar el deterioro de los organismos.
Los muestreos se efectuarán en dos secciones transversales del río Paraná y su llanura aluvial. Una de ellas se ubicará en las proximidades de la ciudad de Reconquista, en el sitio RAMSAR “Jaaukanigás”. La segunda sección transversal se ubicará en el Paraná Inferior, en el sector sur del Parque Nacional Pre-Delta. La recolección de ejemplares de esponjas y de L. fortunei se llevará a cabo en dichas transectas sobre las riberas del cauce principal del río Paraná y de los cauces secundarios presentes en la transecta, en la zona marginal de las lagunas y en la vegetación de la zona de transición acuático-terrestre.
Bibliografia
Darrigran, G. & I.Ezcurra de Drago. 2000. Invasion of the exotic freshwater mussel Limnoperna fortunei (Dunker, 1857) (Bivalvia,Mytilidae) in South America. The Nautilus 114 (2): 69-73.
Ezcurra de Drago, I. 1972. Contributo alla conoscenza delle spugne d'acqua dolce d'Italia (Porifera, Spongillidae). Mem.Ist.Ital.Idrobiol. 29:109-127.
________________ 1975. El género Ephydatia Lamouroux (Porifera, Spongilllidae). Sistemática y distribución. Physis B 34:157-174.
________________ 1976. Ephydatia mülleri Lieberkühn) in Africa, and the systematic postion of Ephydatia japonica (Hilgendorf) (Porifera, Spongillidae), Arnoldia 25: 1-7.
________________ 1988. Nuevos aportes a la taxonomía y distribución de las especies de Heteromeyenia Potts, en Argentina. Stud.Neotrop.Fauna.Envin. 23:137-147.
_______________ 1993. Distribución geográfica de las esponjas Argentinas (Porifera: Spongillidae, Potamolepidae y Metaniidae). Relaciones zoogeográficas y vías de poblamiento, en: Boltovskoy & López (Ed.), Conferencias de Limnología , pág. 115-125, Instituto de Limnología “Dr. Raúl A. Ringuelet”, La Plata, Buenos Aires, Argentina.
________________ 1995. Porifera, en “Ecosistemas de aguas continentales. Metodología para su estudio”. Ed. Dres. G. Tell y E. Lopretto. Ediciones Sur. Argentina.
Harrison, F.W. 1974. Sponges (Porifera:Spongillidae). En: Pollution Ecology of Freshwater Invertebrates. Ed.: C.W.Hart, Jr & S.L.H.Fuller, Academic Press, New York, pp. 29-66.
_____________ 1988. Utilization of freshwater sponges in paleolimnological studies. En: Paleolimnology. Aspects of freshwater palaecology and biogeography . Ed.: J. Gray. Elsevier, Amsterdam, pp. 387-397.
Harrison, F.W. & B.G.Warner. 1986. Fossil freshwater sponges (Porifera, Spongillidae) from Western Canada: An overlook group of quaternary paleocological indicators. Trans.Am.Microsc.Soc.105:110-120.
Jewell, M.E. 1935. An ecological study of the freshwater sponges of northeastern Wisconsin. Ecol.Monogr. 5:462-501.
Manconi, R y R. Pronzato. 2002. Suborder Spongillina subord. Nov.: Freshwater Sponges, en Hooper & Van Soest (Ed.), Sistema Porifera: A Guide to the Classification of Sponges . Kluwer Academic/Plenum Publishers. New York
Morello, J.M. 1984. Perfil ecológico de Sudamérica. Volumen I. Características estructurales de Sudamérica y su relación con espacios semejantes del planeta. Instituto de Cooperación Iberoamericana. Barcelona, 93 pág.
Moore, W.G. 1953. Louisiana freshwater sponges, with ecological observations on certain sponges of the New Orleans area. Trans.Amer.Microsc.Soc. 72:24-32.
Muller, W.E.G., R.K. Zahn y A. Maidhof. 1982. Spongilla gutenbergiana n.sp., ein susswasserschwamm aus dem Mittel-Eozan von Messel. Senckenbergiana Lethaes, 63 (5/6)): 465-472.
Ott, E y W. Volkheimer. 1972. Palaespongilla chubutensis n.g. et n.sp. ein susswasserschwamm aus der kreide Patagoniens. N. Jb. Geol. Palaont. Abh., 140: 49-63.
Pastorino, G., G.Darrigran, S.Martin & L.Lunaschi. 1993. Limnoperna fortunei (Dunker, 1857) (Mytilidae), nuevo bivalvo invasor en aguas del Río de la Plata. Neotropica 39 (101-102):34.
Poirrier, M.A. 1969. Louisiana freshwater sponges: taxonomy, ecology and distribution. Ph.D. Thesis. Louisiana State Univ. Microfilms Inc., Ann Arbor, Michigan, Nº 70-9083.
Racek, A.A. 1966. Spicular remains of freshwater sponges. En: The history of Laguna de Petenxil. Mem.Conn.Acad.Arts.Sci. 17:78-83.
____________ 1970. The Porifera. En: Ianula: An account of the history and development of the Lago di Monterosi, Latium, Italy. Trans.Am.Philos.Soc. 60(pt.4):143-149.
____________ 1974. The waters of Merom: A study of lake Huleh IV. Spicular remains of freshwater sponges (Porifera). Arch.Hydrobiol., 74:137-158.
Racek, A.A. & Harrison, F.W. 1974. The systematic and phylogenetic position on Palaespongilla chubutensis (Porifera, Spongillidae). Proc.Linn.Soc.N.S.W.99:157-165.
Rapoport, E.H. 1968. Algunos problemas biogeográficos del Nuevo Mundo con especial referencia a la región Neotropical. En: Biologie del'Amerique Australe, Vol.IV. Ed. C. Delamare Deboutteville & E.H. Rapoport. CNRS & CNICT. Paris, pp. 54-110.
Ringuelet, R.A. 1961. Rasgos fundamentales de la zoogeografía de la República Argentina . Physis 22:151-170.
Soldano, F.A. 1947. Régimen y aprovechamiento de la red fluvial argentina. Parte I. El río Paraná y sus tributarios. Ed. Cimera, Buenos Aires, 264 pág.
Volkmer-Ribeiro, C. y J. Reitner. 1991. Renewed study of the type material of Palaespongilla chubutensis Ott and Vokheimer (1972). Pp: 121-133, In: J. Reitner y J. Keupp (eds.). Fossil and Recent Sponges . Springer-Verlag, Berlin Heidelberg.
Volkmer-Ribeiro, C. y S.M. Pauls. 2000. Esponjas de agua dulce (Porifera, Demospongiae) de Venezuela. Acta Biológica Venez. , 20(1): 1-28.
Fig. 1. Isolíneas de riqueza específica de esponjas ( por cuadrados de 100 x 100 kilómetros)

Lamina: Esponjas de agua dulce: 1. Drulia browni, laguna Setúbal, Paraná Medio, Santa Fe. 2. Detalle de la trama esquelética y gémulas de D. browni. 3. Trochospongilla delicata, laguna Los Naranjos, Paraná Medio, Santa Fe. 4. Detalle de la trama esquelética y gémulas de T. delicata. 5. Uruguayella repens, Federación, río Uruguay. 6. Detalle de la trama esquelética de U. repens de Federación. 7. Detalle de la trama esquelética y ósculos de U. repens de la laguna Setúbal. 8. Uruguaya corallioides, Federación, río Uruguay. 9. Limnoperna fortunei bisada sobre un ejemplar de D. browni, embalse Salto Grande, Río Uruguay.

Tabla 1. Especies registradas en ambientes de la región Mesopotámica