Aporte al conocimiento de ciclos de materia orgánica (MO) en formaciones boscosas de la llanura de inundación del Río Paraná: Area PNPD
Pamela ZAMBONI y Pablo ACEÑOLAZA1
1CICyTTP-CONICET. Matteri y España (3105) Diamante. Entre Ríos. FCA - UNER - acenolaza@hotmail.com.
pamelazamboni@hotmail.com
Abstract: CONTRIBUTION TO KNOWLEDGE TO ORGANIC MATTER CYCLES (OM) IN FOREST OF THE PARANA RIVER FOODPLAIN .A study was made to know the organic matter (OM) cycling in tree representative forest of Paraná River Floodplain. It was carried out in Pre-Delta National Park (Entre Ríos, Argentina) during the 2001/2 period. The time-course of the litter fall for different organs of the trees was followed, also litter decomposition rates were followed. The mean total production of litter in Salix humboldtiana forest was 475 gr/m 2 /year; 464 gr/m 2 /year for Tessaria integrifolia and 245 gr/m 2 /year for Albizia inundata . The temporal pattern showed maximum fall values during winter-spring period (July-January). S. humboldtiana shows lower litter decomposition rates ( k ) and higher permanence time; on the other hand, A. inundata shows higher decomposition rate and consequently lower permanence as forest litter.
Key words: Organic matter, decomposition rate, wetlands, forest litter, flooding pulse, floodplain, Paraná River.
Palabras claves: Materia Orgánica, Descomposición, Bosques ribereños, Río Paraná.
Introducción
La llanura aluvial del río Paraná se caracteriza por la dinámica y heterogeneidad temporal y espacial de sus ambientes. Su geomorfología está determinada por procesos geológicos, fluviales, climáticos (Iriondo, 1988; Passeggi, 2000).
Los ecosistemas de la región están adaptados a las variaciones ambientales. Su integridad es mantenida, entre otras variables, por la dinámica hidrológica (Junk, et al, 1989; Sparks, 1995). La composición y producción de la vegetación responden a esta dinámica.
La caída de hojas, flores, ramas, frutos, corteza, entre otras estructuras que conforman la hojarasca, representa la vía principal en el flujo de materia y energía al piso forestal (Margalef, 1974; Spurr y Barnes, 1982; Aceñolaza, 1996).
En los ambientes de llanura de inundación los ciclos de materia y energía tienen propiedades propias. La entrada, circulación y salida de MO y nutrientes en el ecosistema presenta relación con el régimen hidrológico (Neiff y Poi de Neiff, 1990). Así los ciclos de MO se ven acelerados, dotando a estas regiones de una alta productividad en comparación con otros ambientes (Brinson et al . 1980, Junk et al . 1989; Sparks, 1995).
El aporte de MO al suelo a lo largo del año está ligado a factores climáticos, fluviales, edáficos, manejos forestales y propios de la vegetación, entre otros. En el presente trabajo se analizó el patrón temporal de aporte de hojarasca y se establecieron relaciones con variables ambientales (altura del río y precipitaciones) durante el período de estudio.
Desde esta perspectiva es poco conocido el comportamiento de los sistemas forestales en la llanura de inundación del Río Paraná, destacándose el trabajo realizado por Neiff y Poi de Neiff (1990) para Tessaria integrifolia . Se seleccionaron tres bosques dominados por Salix humboldtiana (sauce criollo), Tessaria integrifolia (aliso del río) y Albizia inundata (timbó blanco), basándonos en la representatividad areal que estas comunidades poseen en la región.
El objetivo de este trabajo fue analizar el aporte y la descomposición de MO en tres comunidades arbóreas representativas de la llanura de inundación de Río Paraná. Se trabajó con la hipótesis de que existen diferencias inter e intracomunitarias en los procesos asociados al aporte y descomposición de MO. Estas variaciones estarían relacionadas a las características propias de las especies dominantes (historia de vida), y a la respuesta que cada una posee ante eventos cíclicos como las inundaciones.
Materiales y métodos
El trabajo de campo se llevó a cabo en el Parque Nacional Predelta (PNPD) (Lat. S. 32º 03´ 43” Long. O. 60º 38´39”), departamento Diamante, provincia de Entre Ríos (Figura 1).
El clima es templado/cálido y húmedo, las temperaturas medias anuales rondan los 19ºC. El régimen pluviométrico es de 900 mm. al año (Rojas y Salusso, 1987). La vegetación corresponde de acuerdo a Cabrera (1994), a la Región Neotropical Dominio Amazónico (Provincia Paranaense, Selvas Marginales).
Las áreas de muestro se seleccionaron siguiendo el criterio de estratificación por especie dominante (Lewis y Francheschi, 1979). Estos bosques tienen características de monoespecíficos o de baja diversidad; densidades entre 833 ind/ha en el bosque de sauce a 1063 ind/ha en el bosque de timbó (aliso 992 ind/ha); área basal entre 19 a 34 m 2 /ha (sauce 34 m 2 /ha, aliso 30 m 2 /ha y 19 m 2 /ha timbó); y alturas medias de 10 metros en el bosque de sauce, 8 metros en el de aliso y 4.5 metros en el de timbó (Zamboni, 2003).
El estudio del ciclo de MO abarca tanto la producción como la descomposición de la MO producida. Para estimar la producción de hojarasca se utilizó el método de recolección de la hojarasca (Kira y Shidei, 1967) que consiste en cosechar, secar y pesar muestras de material caído en intervalos de tiempo predeterminados. Esta metodología es ampliamente utilizada en el ambiente científico, tanto las características del muestreo y procedimiento analítico empleados han sido profusamente reportados en otros estudios (Shure y Phillips, 1978; Brinson et al , 1980; Neiff y Poi de Neiff, 1990; Santa Regina et al , 1991; Clarke y Allaway, 1996; Williams-Linera y Tolomé, 1996; Grigg y Mulligan, 1999).
El trabajo de campo consistió en la recolección periódica (mensual) de todo el material vegetal caído en cajones de recolección de hojarasca en cada bosque durante el período estudiado. Se utilizaron 6 dispositivos de recogida por bosque, distribuidos aleatoriamente; cada uno de 0.25 m 2 de superficie y 10 cm de alto. Las muestras se colocaron en bolsas de papel y se secaron en estufa a 80ºC hasta llegar a un peso constante. El material se clasificó separando por categoría los fragmentos reconocibles de hojarasca (hojas, ramas, flores y frutos). Esta diferenciación se realizó sobre las especies dominantes de cada bosque. La hojarasca correspondiente a las demás especies (estratos arbustivo, herbáceo) y fragmentos no reconocibles se agruparon en una categoría denominada “varios”. Se analizó el patrón temporal del aporte de hojarasca para cada bosque y para la especie dominante de cada bosque.
Los datos fueron analizados estadísticamente. Se obtuvieron medias, desviaciones estándar e intervalos de confianza (IC) (alfa de 0.05) para cada mes y bosque.
Se efectuaron análisis de homosedasticidad y normalidad de los datos. Se examinó la existencia de diferencias estadísticamente significativas entre bosques y meses mediante el test no paramétrico de Kruskal Wallis.
Para estimar la velocidad de descomposición de hojarasca, se utilizó la metodología propuesta por Jenny et al. (1949), que consiste en la recolección de muestras del mantillo a intervalos regulares de 2 meses durante el año de estudio. El tratamiento en laboratorio de las muestras fue similar a las de aporte. Se registró el peso utilizando una balanza de precisión. Con los valores obtenidos se estimó la velocidad de descomposición aplicando el coeficiente de descomposición de hojarasca “ k ” (constante de Jenny) y el tiempo medio de permanencia de residuos (constante Olson).
Resultados y discusiones
Los resultados se presentan en forma de tabla y gráficos.
El aporte anual de hojarasca varió en los tres bosques entre 243 gr/m 2 en el bosque de timbó y 475 gr/m 2 en el de sauce (Tabla 1). No se observaron diferencias significativas en el aporte anual de hojarasca entre los bosques de sauce y aliso, aunque si con el de timbó ( p <0.05).
Al examinar la hojarasca aportada por la especie dominante (en el caso del bosque de sauce es Salix humboldtiana , en el de aliso es Tessaria integrifolia y en el de timbó es Albizia inundata ), se pudieron observar diferencias significativas ( p <0.05) entre las tres especies estudiadas. Esto, junto con lo comentado en el párrafo anterior, podría indicar que la contribución de otros estratos de las comunidades (otras especies arbóreas y sotobosque) se constituirían como un factor homogeneizador, nivelando el ingreso de MO dentro a cada comunidad.
El aporte total anual de hojarasca en los bosques estudiados es algo menor a la media universal de 5600 kg/ha/año calculada para bosques templados cálidos (Brinson et al , 1980). Neiff y Poi Neiff (1990) registraron valores de 815 gr/m 2 para bosques de Tessaria integrifolia , este valor es superior al estimado en el presente estudio. Cabe destacar que para nuestro estudio, la densidad de individuos por hectárea es 8 veces menor que el encontrado por Neiff y Poi Neiff (1990), de lo que podría inferirse una posible relación entre densidad o edad sucesional del bosque y el aporte de órganos caducos. Grigg y Mulligan (1999) afirman que altos valores de densidad de arbolado pueden explicar altos valores en el aporte de hojarasca, mientras que Aceñolaza (1996) encontró una estrecha relación entre el aporte y la edad sucesional de bosques de Alnus acuminata .
En cuanto a la composición porcentual de dicho aporte, las hojas contribuyeron en el bosque de sauce con un 63%, valor similar al supuesto promedio universal de 70% (Shure y Phillips, 1987), ramas 20%, flores y frutos 2% y “varios” con un 15%. En el bosque de aliso las hojas constituyeron el 36% del total anual, ramas 10%, flores y frutos 2% y “varios” el 52%. En el bosque de timbó el porcentual del aporte se distribuyó de la siguiente manera, hojas con el 23 %, ramas 19%, flores y frutos 2% y “varios” con el 56% (Zamboni, 2003).
En términos relativos la contribución de flores y frutos es de 2% (categoría con menor representación) y la de ramas 10-20% en los tres bosques. “Varios” se constituyó como la categoría más representada en los bosques de aliso y timbó, igual que hojas en el bosque de sauce.
El patrón temporal del aporte en los tres bosques, evidencia una cronología estacional con un periodo máximo en los meses de julio a enero (70% del aporte anual; p < 0.05 para sauce y aliso). Observándose un pico en el mes de mayo, correspondiente a la caída de hojas de otoño (Figura 2 a, b y c).
Por el contrario, si bien el período de máximo aporte de hojarasca para Albizia inundata se registró en los meses de abril a septiembre (Fig. 2 a), no se observaron diferencias significativas ( p >0.05) en el patrón temporal del aporte (Tabla 1). Podemos hipotetizar que Albizia inundata no define el patrón temporal del aporte en su bosque; posiblemente son las especies que componen su sotobosque las que explican el modelo de aporte para el bosque.
La curva de aporte de hojarasca del bosque de sauce y aliso (Fig. 2 b y c) copia el modelo de aporte sus especies dominantes.
Neiff y Poi de Neiff (1990) encontraron para bosques de aliso que los meses de mayor aporte de hojarasca coincidían con el período de déficit hídrico (octubre a enero) concordando los registros más bajos con los pulsos de inundación (otoño). Dichos patrones de aporte son similares a los observados en el presente estudio donde la caída de hojarasca exhibe altos valores en los meses de primavera y verano, con la llegada de aumento de temperatura y radiación, período que en esta región coincide con bajo nivel hidrométrico del río. Sin embargo esta relación no es estadísticamente clara para toda la secuencia anual de datos, ya que el coeficiente de determinación (R 2 ) entre la variables (altura del río y aporte) resultó menor a 0.50 para los tres bosques. El mismo resultado se obtuvo al realizar las regresiones entre precipitaciones y aporte; por lo que se estima que variables inherentes a las características propias de las especies (factores genético/evolutivos, ecológicos, etc) contribuirían, conjuntamente con las anteriores variables, a explicar este modelo de aporte.
La velocidad de descomposición de hojarasca está inversamente relacionada con el tiempo medio de permanencia de residuos, los resultados obtenidos permiten reafirmar que la hojarasca acumulada en el bosque de sauce se degrada a una tasa menor que en los otros bosques, mientras que la del bosque de timbó es la de menor permanencia (y mayor velocidad de descomposición). En el bosque de timbó, la tasa de circulación de MO (y por lo tanto nutrientes) es más rápida que en los otros bosques. Estas diferencias pueden atribuirse a características especificas propias del material en proceso de descomposición (contenido de lignina, N, P, etc.).
Conclusiones
Se analizó el aporte y la descomposición de MO proveniente de la hojarasca en tres bosques representativos de la llanura de inundación del Río Paraná. Se encontraron diferencias significativas en el aporte total anual entre los bosques de sauce y aliso contra el de timbó. El aporte de MO al sustrato es de 4750 kg/ha/año para el bosque de sauce, de 4640 kg/ha/año para el de aliso y de 2432 kg/ha/año en el de timbó. Las estructuras de las plantas (hojas, ramas, flores y frutos) exhibieron proporciones diferentes de aporte.
Se observó un modelo estacional de aporte, con aproximadamente el 70% del flujo anual de MO en los meses de julio a enero en los tres bosques. El período de mayor aporte de hojarasca se observó en primavera, ligado al aumento de temperatura, radiación, estrés hídrico, entre otros factores. Idénticas conclusiones han sido reportadas por Neiff y Poi de Neiff (1990) y Grigg y Mulligan (1999).
El patrón temporal del aporte de la especie dominante presenta una curva similar al de la hojarasca en los bosques de aliso y sauce, mientras que en el bosque de timbó el período de mayor aporte de la especie dominante se registró en los meses de abril a septiembre.
La velocidad de descomposición de hojarasca es diferente en los tres bosques, siendo el bosque de sauce el que presenta la menor tasa de descomposición y el bosque de timbó el de mayor. En este ultimo bosque la acumulación de MO en el sustrato es menor, tanto por su menor aporte como su mayor tasa de descomposición.
Agradecimientos: A la Administración de Parques Nacionales, a su personal en el PNPD, Reynaldo Zanello, Miguel Dedeck, Omar Bejarano. Al PTC Raúl D´Angelo por la colaboración en trabajos de campo. Parcialmente financiado por ANPCyT, PICT 4006/98 y PID-UNER 2089/1.
Bibliografía
Aceñolaza. P. 1996. Estructura y dinámica de bosques de aliso (Alnus acuminata HBK spp acuminata) de la provincia de Tucumán . Tesis doctoral. Universidad Nacional de Tucumán. pp 286.
Brinson, M. M.; H.D Bradshaw; R. N. Holmes y J. Elkins. 1980. Litterfall, stemflow and throughfall nutrient fluxes in an alluvial swamp forest. Ecology . 61(4), 827-835.
Cabrera, A. L. 1994. Regiones fitogeográficas Argentinas. Enciclopedia Argentina de Agricultura y Jardinería (1). Ed. ACME Buenos Aires. 298 pp
Clarke, P. J.y W. G. Allaway. 1996. Litterfall in Casuarina glauca Coastal Wetland Forest. Australian Journal of Botany 44, 373-380.
Grigg, A. H. y D. R Mulligan. 1999. Litterfall from two eucalypt woodlands in central Queensland. Australian journal of Ecology. 24, 662-664.
Iriondo, M. 1988. Llanura Aluvial del Río Paraná. Museo de Ciencias Naturales y Antropológicas Profesor Antonio Serrano. Guía de campo Nº 4 . Paraná.
Jenny H., S.P. Gessel y F.T. Bingham, 1949. Comparative study of decomposition rates of organic matter in temperate and tropical regions. Soil Science 68: 419-432.
Junk W.; P. Bayley y R. Sparks. 1989. The flood pulse concept in river-floodplain systems. Fishand Aquatic Science 106: 110-127.
Kira, T y T. Shidei. 1967. Primary productivity and turnover of organic matter in different forest ecosystems of the western Pacific . Japan Journal Ecology. 17: 70-87.
Lewis J. J. y E. A. Francheschi. 1979. Notas sobre la dinámica de la vegetación del valle del río Paraná. ECOSUR . 6:145-163.
Margalef, R. 1974. Ecología . Ed. Omega. España. 951 pp.
Neiff J. J. y A. Poi de Neiff. 1990. Litterfall, leaf decomposition and litter colonization of Tessaria integrifolia (compositae) in the Paraná river floodplain. Hidrobiología. 203: 45-52. Belgium.
Passeggi E. 2000. Caracterización sedimentológica del material parental de los suelos asociados a los depósitos de cauce del tramo medio del río Paraná . Tesis doctoral. Universidad Católica de Santa Fé. 216 pp.
Rojas, A.E. y J. H. Salusso. 1987. Informe climático de la provincia de Entre Ríos . Publicación técnica nº 14. INTA. Paraná. 20 p.
Santa Regina, I.; J. Gallardo; M. Rico; A. Martin; H. A. Gallego; G. Moreno y S. Cuadrado. 1991. Datos preliminares sobre biomasa aérea, producción y características edafoclimáticas de ecosistemas forestales de Quercus pyrenaica (Sierra de Gata, Salamanca). Studia Ecologica 8: 147-158.
Shure, D.J. y D.L Phillips. 1987. Litter Fall Patterns within Different-sized Disturbance Patches in a Southern Appalachian Mountain Forest. The American Midland Naturalist . 348-357.
Sparks, R. E.. 1995. Need for Ecosystem Management of Large Rivers and their Floodplains. Bio Science . 45: 168-182.
Spurr S. H. y B. V. Barnes. 1982. Ecología forestal . Ed. AGT. México. 690 pp.
Williams-Linera G. y Tolome J. 1996. Litterfall, Temperate and Tropical Dominant Trees, and Climate in a Mexican Lower Montane Forest. Biotrópica 28: 649-656.
Zamboni, L.P. 2003. Dinámica de materia orgánica en tres bosques de la llanura de inundación del Río Paraná . Tesina de Licenciatura en Biodiversidad. FHC-UNL. 69 pp. Santa Fe.
Fig. 1. Mapa general y de ubicación del área de estudio.
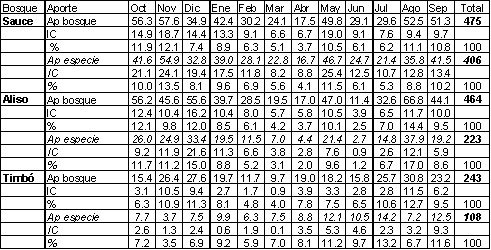
Tabla 1. patrón temporal del aporte total de hojarasca para el bosque (Ap bosque) y para la especie dominante (Ap especie). Los resultados se presentan en valores absolutos (gr/m2) y en porcentajes. IC: intervalo de confianza (alfa= 0.05). Período octubre 2001 a septiembre 2002. Se destacan los valores anuales.
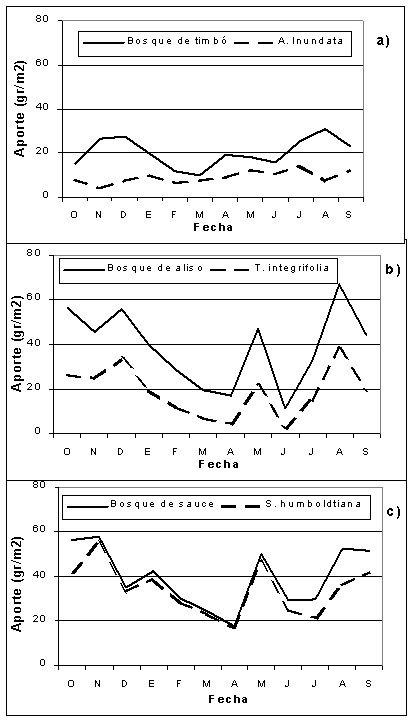
Fig. 2. Patrón temporal de aporte de hojarasca, en gr/m2, del bosque y de la especie dominante en cada bosque; a) bosque de timbó, b) bosque de aliso y c) bosque de sauce. Período de estudio: octubre 2001-septiembre 2002.