Las especies exóticas invasoras en los sistemas de humedales. El caso del Delta inferior del Río Paraná
Fabio A. KALESNIK1y2 y Ana Inés MALVAREZ2
1CICYTTP. Centro de Investigaciones Científicas y de Transferencia de Tecnología a la Producción. CONICET. Argentina.
2Grupo de Investigación en Ecología de Humedales. Dpto. Ecología, Genética y Evolución. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires.
Abstract: EXOTIC INVADER SPECIES IN WETLAND ECOSYSTEMS. THE CASE OF THE LOW DELTA OF THE PARANA RIVER.-. Wetlands are ecosystems whose functioning relies on hydrologic regimes and small variations of flooding pulses or flooding levels that may produce massive changes in the local biota. These systems are particularly vulnerable to invasion processes, where variations of hydrologic regimens may cause changes in community composition and structure, and are considered to be one of the causes of incorporation of alien species.
The delta of the Paraná river may be considered as a mosaic of wetlands, where 11 landscape units may be characterized by different flooding regimes and landscape patterns. In particular, the “Lower Delta Islands” Unit, is characterized by a highly fragmented landscape constituted by natural (marsh and relict patches of native forests) and anthropic environments ( Salix commercial afforestations). These juxtapositions may increase considerably the probability of dispersion of alien species.
Burkart (1957) mentions 29 exotic species for natural environments. In more recent works only 11 exotic species are mentioned. Lowland environments (marsh) present 4 exotic species (iris, Iris pseudacorus ; honeysuckle, Lonicera japónica ; Chinese privet, Ligustrum sinense and blackberry, Rubus spp. ) but these species do not behave as invaders, since they present very low constancy and cover values. On the other hand, relict patches of native forests, present high cover values for privet, ( Ligustrum lucidum ), honeysuckle, White mulberry ( Morus alba ), and maple ( Acer negundo ).
What respects anthropic environments, Valli (1990) analyzed succesional trends in lowland forestations, mentioning 5 exotic species (iris, Faux indigo ( Amorpha fructicosa ), honeysuckle, and privet), all of them with low cover values. Kalesnik (2001) studied commercial forestations on levee that presented 16 exotic species, of which only 7 behaved as invaders: privet, Fraxinus pennsylvanica , black acacia ( Gleditsia triacanthos ), honeysuckle, blackberry, maple and Ligustrum lucidum . The development of each species varied among different subunits (“A”, “B” or “C”) of the Lower Delta. While unit “A” presented a very low invasion index, low cover development of alien, and none of the species behaving as invaders, the other two units (“B” and “C”) presented mean values of the invasion index and a high cover of the mentioned exotic species. In unit “A”, the fluvial influence of the Paraná river, acting together with extraordinary flooding events and the fire caused intentionally for hunting activities, may be a possible explication for the absence of invader species.
Finally, it may be stated that of a total of 102 exotic species introduces in the Lower Delta, 8,82% behave as invaders in the island sector. This value is in concordance with the pattern found for temperate ecosystems, where only a 10% of introduced species behave as invaders, in contrast to tropical ecosystems, where that proportion is close to 100%.
Key words: wetlands, Low Delta of the Paraná river, exotic and invader species
Palabras claves: Humedales, Bajo Delta del Río Paraná, especies exóticas e invasoras
Las especies exóticas invasoras
Una determinada especie puede ser considerada como introducida (exótica) cuando la misma ha sido intencionalmente o accidentalmente transportada por el hombre a un área fuera de su rango de distribución geográfica natural y como una especie invasora , cuando la misma , una vez introducida puede expandir su población (o rango de distribución) en la nueva situación geográfica sin necesidad de la intervención humana. (Usher 1991; Pysek, 1995).
En la bibliografía también es muy frecuente la utilización del término maleza (weed), indicando con el mismo, a aquellas especies que crecen en áreas disturbadas por el hombre, independientemente del uso al que estén sometidas las mismas y el origen biogeográfico de las especies consideradas (Baker, 1965). Otros autores también utilizan el término maleza para definir las especies pestes de sistemas agrícolas y otros sistemas manejables que puedan llegar a reducir el valor económico de los mismos (Bazzaz. F. A, 1986).
Debido al múltiple uso utilizado en la definición de este último término, nosotros trabajaremos con los términos de especies introducidas e invasoras ya que nos parecen términos más precisos para un entendimiento ecológico de la problemática planteada.
Ambientes susceptibles a los procesos de invasión
“Muchas comunidades naturales han sido o están siendo invadidas por especies exóticas, las cuales afectan la estructura y procesos que se desarrollan en los ecosistemas en que se encuentran las mismas” (Rejmánek, 1995).
En general, los procesos invasivos han sido asociados a áreas que han sido disturbadas en alguna medida por la acción del hombre, considerando al mismo, como el principal agente de introducción de especies invasoras en los distintos ecosistemas. (Baker, 1974).
En este sentido, la intervención antrópica podría actuar como un disturbio creador de parches en la vegetación existente, a partir del cual se incrementaría la disponibilidad de recursos necesarios para el establecimiento e incorporación de las especies invasoras al sistema. (Pickett and White, 1985; Macdonald et al ., 1988).
Esta última interpretación, sobre la relación obligada de disturbio de origen antrópico-invasión fue puesta en discusión en los últimos años. En este sentido, diversos trabajos realizados en ambientes naturales de reservas tropicales caracterizados por no presentar registros históricos de acción antrópica, mostraron situaciones que iban desde niveles bajos a altos de invasión de especies vegetales, pudiendo plantear con ello que “ninguna región del planeta podría llegar a estar libre de especies invasoras”. (Usher, M. B, 1991; Cowie y Werner, 1993).
Debido a la susceptibilidad de los diversos ecosistemas a los procesos invasivos y a las modificaciones que éstos producen en los mismos, numerosos trabajos plantean la necesidad del control o eliminación de las principales especies invasoras (Usher, 1991; Cowie y Werner, 1993, entre muchos otros).
Sin embargo, existen otros trabajos que interpretan a los procesos invasivos simplemente como una parte de los procesos normales de las distintas comunidades (Diamond y Case, 1986; Chesson, 1986), resultando interesante el planteo de Hobbs (1989), al considerar que las actividades humanas serían el principal factor de aporte de especies invasoras que pasarían a formar parte de las comunidades “naturales”. A su vez, existirían situaciones en las cuales las especies invasoras una vez incorporadas al sistema, podrían recrear las condiciones ambientales favorables para el crecimiento de las especies nativas en ecosistemas degradados antrópicamente (De Pietri, 1991).
Procesos invasivos en ambientes de humedales
Los humedales son ecosistemas cuyo funcionamiento depende del régimen hidrológico y pequeñas variaciones en el pulso de inundación o en los niveles de anegamiento pueden producir cambios masivos en la biota presente (Mitsch y Gosselink, 2000).
Estos tipos de ambientes son particularmente susceptibles a los procesos de invasión y las variaciones en el régimen hidrológico pueden causar modificaciones en la composición y estructura de las comunidades y son consideradas como una de las causas de incorporación de especies invasoras.
Howe and Knopf (1991), observaron que la variación en el régimen hidrológico fue la principal causante para el éxito de la invasión de dos especies arbóreas en los humedales riparios del oeste árido de los Estados Unidos ( Eleagnus angustifolia , olivo ruso y Tamarix chinensis ), e incluso el motivo de la pérdida de una especie arbórea nativa ( Populus fremontii ). A su vez, el grado de invasión de estas especies fue de tal magnitud que llegó a modificar los procesos sucesionales de los bosques riparios de la región. (Brinson et al. 1981).
Como otro ejemplo de lo mencionado anteriormente, podemos citar el trabajo de Beerling, 1991 en el cual se describe el proceso de invasión en humedales riparios de Gran Bretaña. En el mismo, se pone de manifiesto la importancia de los eventos de inundación como un proceso clave en el éxito de dispersión de los rizomas de Reynoutria japonica mediante los cuales invade exitosamente los distintos ambientes. De este modo, también queda claro que cualquier modificación en estos eventos de inundación podría influir sobre la dinámica del proceso invasivo.
Otros autores, relacionan los procesos invasivos con los incrementos de nutrientes que se pueden producir en una comunidad (Hobbs y Atkins, 1988), mientras que otros autores también relacionan la simplicidad estructural que caracterizan a algunos tipos de humedales con el éxito invasivo de algunas especies (Taylor & Dunlop, 1985). Como ejemplo de esto último, se puede mencionar un trabajo realizado en el Kakadu National Park (Australia), en el cual grandes extensiones de humedales dominados por pastos y juncos fueron invadidos por Mimosa pigra (arbusto americano-africano tropical) convirtiendo a los mismos en grandes arbustales (Braithwaite et al . 1989). En el mismo, se plantea que la simplicidad estructural de la comunidad natural asociada a condiciones de disturbio estarían favoreciendo el proceso invasivo (Cowie y Werner, 1993).
Por otro lado, los sistemas de humedales están siendo sometidos a un intenso manejo antrópico, con lo cual a nivel de paisaje, se produce una elevada fragmentación del los mismos que conduce a la yuxtaposición de ambientes naturales y antropizados, como así también a la modificación de sus principales variables condicionantes. Este tipo de situaciones incrementaría en forma considerable la probabilidad de dispersión de especies invasoras dentro de los ambientes naturales (Hobbs. R. J, 1989).
Las especies exóticas invasoras en el Bajo Delta del Río Paraná
La región del Delta del Río Paraná puede ser considerada como un macromosaico de humedales en el cual se encuentran 11 unidades de paisaje caracterizadas por diferentes regímenes de inundación y patrones de paisaje. En particular, analizando la unidad del sector de islas del Bajo Delta, se puede mensionar un paisaje altamente fragmentado constituido por ambientes naturales (pajonales y parches relictuales de bosque nativo) y por ambientes atropizados (forestaciones comerciales de salicáceas). Esta yuxtaposición de ambientes incrementaría en forma considerable la probabilidad de dispersión de especies invasoras.
Del total de las 632 especies vegetales citadas para el Delta Inferior (Kalesnik y Malvárez, 1996) el 16.14 % es de origen exótico (102 especies). Cabe aclarar que dicha región abarca tanto el sector de islas propiamente dicho, así como sectores de la planicie aluvial situados en la provincia de Entre Ríos.
El porcentaje de especies introducidas coincide con los patrones de invasión encontrados en distintos sistemas templados y reservas naturales tropicales analizados a nivel mundial, en los cuales se encontró la existencia de especies exóticas en una proporción cercana a valores del 10 % en relación a la flora local (Macdonald y Frame, 1988; Cowie y Werner. P. A., 1993).
En cuanto al origen, más de la mitad de estas especies introducidas son originarias de Europa (63.70 %) y en menor proporción de Asia (17.64 %), América del Norte (8.82 %), Africa (5.88 %) y Oceanía (3.92 %).
A su vez, las mismas están representadas por 33 familias vegetales, pudiéndose destacar las siguientes: Asteraceae (21 especies), Fabaceae (10 especies), Poaceae (9 especies), Brassicaceae (6 especies), Polygonaceae (5 especies) y las familias restantes con menos de 4 especies cada una.
Las tres familias más representadas en cuanto al número de especies exóticas coinciden con las tres familias más representadas de la flora del Delta Inferior (Kalesnik y Malvárez, 1996) y de la flora de la región Metropolitana de Buenos Aires (Matteucci et al ., 1999). Por otro lado, las cuatro primeras familias mencionadas coinciden con el patrón mencionado por Pysek (1998). En dicho trabajo, del análisis de veintiséis floras representativas de distintos hábitats en todo el globo se plantea que las familias mencionadas poseen el mayor número de especies exóticas a nivel de la flora local.
En particular, en el sector de islas del Bajo Delta se puede mencionar la presencia de un grupo de 29 especies exóticas citadas por Burkart en 1957. En trabajos más recientes Kandus (1997) y Kalesnik (2001) encontraron 11 especies exóticas analizando distintos tipos de ambientes naturales. Los ambientes de bajo (pajonales) presentaron sólo 4 especies exóticas (lirio, Iris pseudacorus ; madreselva, Lonicera japónica ; ligustrina, Ligustrum sinense y zarzamora, Rubus spp. ) que no se comportan como especies invasoras ya que presentaron valores muy bajos de constancia y cobertura. Los parches de los bosques nativos de albardón, (como por ejemplo: el seibal y el monte blanco, ) presentaron altos valores de cobertura de algunas de las especies consideradas, destacándose la ligustrina, ligustro ( Ligustrum lucidum ), madreselva, mora ( Morus alba ) y arce ( Acer negundo ). (Kalesnik, 2001).
En relación a los ambientes modificados antrópicamente (neoecosistemas), se destacan los trabajos realizados por Valli (1990) y Kalesnik (2001).
En el primer caso la autora analiza los principales neoecosistemas de ambientes de bajo (forestaciones de Salix spp. ) y reconstruye la tendencia sucesional en relación a los distintos grado de abandono que presentan los mismos. Una de sus principales conclusiones es el reemplazo de especies terrestres por especies adaptadas a condiciones de inundación temporales o permanentes, observando la regeneración del pajonal original a través del tiempo. En dicha tendencia sucesional, se observó la presencia de cinco especies exóticas que alcanzan bajos valores de cobertura. Se destaca la presencia del lirio en las forestaciones con bajo abandono y del falso índigo ( Amorpha fructicosa ) en las forestaciones con alto abandono. La madreselva y ligustrina estuvieron presentes en todos los tipos de neoecosistemas.
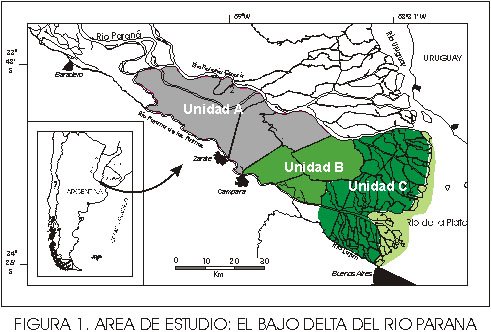
En el segundo caso, los neoecosistemas de albardón presentaron 16 especies exóticas comportándose solamente 7 de ellas como especies invasoras: ligustrina, fresno ( Fraxinus pennsylvanica ), acacia negra ( Gleditsia triacanthos ), madreselva, zarzamora, arce y ligustro. El desarrollo de las mismas presentó variaciones en función de la unidad del Bajo Delta considerada (Figura 1). Mientras que la unidad A, presentó un muy bajo índice de invasión (número de especies exóticas/número de especies totales), un bajo desarrollo de la cobertura vegetal de las especies exóticas y no presentó especies que se comportaran como invasoras, las otras dos unidades (B y C) presentaron valores medios en el índice de invasión y un gran desarrollo de la cobertura de las especies exóticas invasoras mencionadas anteriormente.
La acción conjunta de tres variables (acción fluvial del Río Paraná, eventos de inundaciones extraordinarias y disturbios de orígen antrópico debido a la acción del fuego por causas cinegéticas) y una mayor influencia de las mismas podrían ser una posible explicación de la ausencia de especies invasoras en la unidad A del área de estudio. Bajo este conjunto de condiciones limitantes las especies exóticas sólo se instalarían pero no llegarían a transformarse en especies invasoras. Este concepto cobraría mayor robustez al observar que en los distintos trabajos realizados tanto en los sectores fluviales del Delta Medio y Superior, así como en sectores fluviales aguas arriba en el Paraná Medio no se observó la presencia de las especies exóticas invasoras (Lewis et al. , 1987; Franceschi y Lewis, 1991; Malvárez, 1997).
De este modo, estos resultados coincidirían con lo observado por McIntyre (1988) quien plantea que altos niveles de disturbio endógeno ayudarían a conferir resistencia a la invasión de especies.
En las unidades B y C, las especies vegetales que se desarrollan están asociadas a una menor influencia fluvial del río Paraná y a una mayor influencia del régimen de mareas del Río de la Plata. En las mismas, los picos de crecidas estacionales y extraordinarias del río Paraná se observan en forma amortiguada y los disturbios antrópicos (fuego y ganadería) son de menor magnitud en relación a lo observado en la unidad A. A su vez, el flujo bidireccional producto del régimen de mareas del Río de la Plata, incorporaría un aporte hídrico adicional que compensaría la fase de sequía estacional que caracteriza al régimen hidrológico del río Paraná.
De este modo, las principales especies exóticas que invaden ambas unidades presentarían distintos tipos de adaptaciones que les permitiría desarrollarse bajo las condiciones ambientales descriptas anteriormente.
Esta conclusión reforzaría la tendencia observada en los numerosos trabajos realizados en distintos sistemas a lo largo de la Ribera del Río de la Plata (Dascanio et al 1994; Cagnoni et al 1996; Matteucci et al., 1999; Kalesnik, 2000). En los mismos se menciona la invasión de distintos tipos de ambientes en los cuales se destacan las siete especies exóticas que también invadieron los neoecosistemas de albardón en el Bajo Delta del río Paraná.
Por último, es interesante mencionar que en la unidad B y C las especies invasoras constituyen un nuevo tipo de bosques secundario o “neoecosistema” luego del abandono de las actividades forestales en los albardones de las islas. Los distintos estratos arbóreos y el banco de semillas están dominados por las especies exóticas mencionadas, mientras que las especies arbóreas nativas están presentes sólo con bajos valores de cobertura y densidad (Kalesnik, 2001).
En resumen, una vez analizada la introducción de las especies exóticas en el Delta Inferior y la introducción e invasión de especies exóticas en el sector de islas del mismo, se puede plantear que del total de las 102 especies exóticas introducidas en el Delta Inferior el 8.82 % se comportaron como especies invasoras en el sector de islas.
Esta porcentaje es producto de considerar el conjunto de las 9 especies mencionadas en los resultados de los trabajos de Valli (1990), Kandus (1997) y Kalesnik (2001).
Dicho rango se ubica entre los valores encontrados para ecosistemas templados, en los cuales sólo el 10 % de las especies exóticas introducidas se comportan como invasoras, a diferencia de los ecosistemas tropicales en los cuales dicha proporción es cercana al 100% (Williamson and Brown, 1986; Usher, 1991).
Este porcentaje varía considerablemente si se tiene en cuenta la relación entre especies introducidas y especies invasoras en los distintos tipos de ambientes que caracterizan a las islas del Bajo Delta.
En este sentido, en los neoecosistemas de albardón analizados en el presente trabajo se plantea una relación cercana al 50 %, ya que de las 16 especies exóticas introducidas casi la mitad fueron consideradas especies invasoras.
Los parches relictuales de bosques naturales presentan una relación similar a la anterior ya que 5 de las 11 especies exóticas introducidas podrían ser consideradas como especies invasoras en los bosques analizados de la unidad C.
En relación a los ambientes de bajo, considerando los ambientes naturales y antropizados, se podría plantear que ninguna de las cinco especies exóticas introducidas se comportaron como invasoras a nivel regional.
Bibliografía
Baker, H. G. (1965). Characteristics and modes of origin of weeds. En: Baker, H. G; Stebbins GL (eds.). The genetics of colonizing Species.Academic Press, New York, pp 147-172.
Baker, H. G. (1974). The evolution of weeds. Annual Review of Ecology and Systematics 5: 1-24.
Bazzaz, F. A. (1986). Life History of colonizing Plants: Some Demographic, Genetic, and Physiological Features. En: Ecology of Biological Invasions of North America and Hawaii. Mooney,H. A; J. Drake (eds). pp 96-110.
Beerling, D. J. (1991). The effect of riparian land use on the ocurrence and abundance of Japanese knotweed Reynoutria japonica on selected rivers in South Wales. Biological Conservation 55: 329-37.
Braithwaite, R. W, Lonsdale, W. y Estbergs, J. A. (1989). Alien vegetation and native biota: the impact of Mimosa nigra . Biological Conservation, 48, 189-210.
Brinson, M. M., A. M. Lugo y S. Brown. (1981). Primary productivity, decomposition and consumer activity in freshwater wetlands, Annual Review of Ecology and Systematics 12: 123-161.
Burkart, A. (1957). Ojeada sinóptica sobre la vegetación del Delta del Río Paraná. Darwiniana, 11: 457-561.
Cagnoni, M., Faggi, a. & Ribichich, a. (1996). La vegetación de la Reserva “El Destino”. Parodiana. 9(1-2): 25-44 .
Chesson, P. L. (1986).Environmental variation and the coexistence of species. En: Diamond, J y Case T (eds.). Community Ecology, pp. 240-56. Harper and Row, New York.
Cowie, I.D &. Werner, P.A. (1993). Alien Park species invasive in Kakadu National Park. Australia. Biological Conservation . 63, 127-135 .
Dascanio, L.M., Barrera, M & Frangi, J. (1994). Biomass structure and dry matter dynamic of subtropical alluvial and exotic Ligustrum forest at the Río de la Plata, Argentina. Vegetatio . 115: 61-76.
De Pietri, D.E. (1992). Alien shrubs in a national park: can they help in the recovery of natural degraded forest. Biological conservation. 62, 127-130.
Diamond, J. & Case, T.J. (1986). Community Ecology. Harper & Row, Publishers, New York.
Franceschi, E. A. y J. P. Lewis. (1991). Eaely stages in the Paraná River tall grassland recovery after an extraordinary flood. COENOSES. Vol. 6 (1), pp 47-52.
Hobbs, R. J y Atkins, L. (1988). Effects of disturbance and nutrient addition on native and introduced annuals in plant communities in the Western Australian wheatbelt. Australian Journal Ecology, 13, 43-57.
Hobbs, R.J. (1989). The Nature and Effects of Disturbance Relative to Invasions. En “Biological Invasions: a Global Perspective ”. Drake et al. (eds). SCOPE, pp. 389-405.
Howe, W y F. Knopf. (1991). On the imminent decline of Rio Grande cottonwoods in Central New Mexico. Southwestern Naturalist 36: 218-24.
Kalesnik, F. & Malvárez, A.I. (1996). “Relación entre especies leñosas exóticas y la heterogeneidad ambiental a nivel regional en el Bajo Delta del Río Paraná”. (inédito). Buenos Aires. Argentina.
Kalesnik, F. A; M. Cagnoni; P. Bertolini; R. Quintana, N. Madanes y Malvárez. (2000). La vegetación del “Refugio Educativo de la Ribera Norte”. Invasión de especies exóticas. En prensa. Physis. Argentina.
Kalesnik, F. (2001). Relacion entre las comunidades vegetales de los neoecosistemas de albardon y la heterogeneidad ambiental del Bajo Delta del rio Paraná. Tendencias sucesionales y proyecciones sobre la composicion futura. Tesis Doctoral. Universidad de Buenos Aires.
Kandus, P. (1997). Análisis de patrones de vegetación a escala regional en el Bajo Delta Bonaerense del Río Paraná (Argentina). Tesis Doctoral. Universidad de Buenos Aires.
Lewis, J. P; E. A. Franceschi y D. E. Prado. (1987). Effects of extraordinary floods on the dynamics of tall grasslands of the river Paraná valley. Phytocoenología. Vol. 15 (2), pp 235-251. Berlín. Alemania.
Macdonald, I.A.W. & Frame,G.W. (1988). The invasion of introduced species into nature reserves in tropical savannas and dry woodlands. Biological Conservation 44, 67-93.
Macdonald, I.A.W., Graber, D.M., De Benedetti, S. & Fuentes, E.R. (1988). Introduced species in nature reserves in mediterranean-type climayic regions of the world. Biological Conservation, 44, 37-66.
Malvárez, A.I. (1997). Las comunidades vegetales del Delta del Río Paraná. Su relación con factores ambientales y patrones de paisaje. Tesis Doctoral, Universidad de Buenos Aires, Buenos Aires.
Matteucci, S; J. Morello; A. Rodriguez; G. Buzai y Baxendale, C. (1999). El crecimiento de la metrópoli y los cambios de biodiversidad: el caso de Buenos Aires. En: Biodiversidad y uso dela tierra. Conceptos y ejemplos de Latinoamérica, 580 pp. EUDEBA. Argentina.
McIntyre, S., Ladiges, P. y Adams, G. (1988). Plant species-richness and invasion by exotics in relation to disturbance of wetland communities on the Riverine Plain, NSW. Australian Journal of Ecology 13: 361-373 .
Mitsch, W. & Gosselink, J. (2000). Wetlands . Third Edition. Van Nostrand, New York, 722p.
Pickett, S. T. A. y P. S. White. (1985). The Ecology of Natural Disturbance and Patch Dynamics. Academic Press. New York.
Pysek, P. (1995). On the terminology used in plant invasion studies. En Plant Invasions: General Aspects and Special Problems. Pysek P. et al. (Eds.) SPB. Amsterdam, pp. 71-84.
Pysek, P. (1998). Is there a taxonomic pattern to plant invasions?. Oikos 82: 282-294.
Rejmanek, M. (1995). What makes species invasive?. En Plant Invasions: General Aspects and Special Problems . Pysek P. et al ., (Eds.) . SPB. Amsterdam, pp. 1-3 .
Taylor, J. A y Dunlop, C. R. (1985). Plant communities of the wet-dry tropics of Australia: the Alligator Rivers region. Procceding Ecology Society Australian 13, 83-127.
Usher, M. B., (1991). Biological Invasions into Tropical Nature Reserves. En Ecology of Biological Invasions in the Tropics. Ramakrishnan (Eds.).21-34. 1991.
Valli, S. (1990). Tendencia de las forestaciones en el Delta del Río Paraná y sus implicancias ecológicas. En Condicionantes ambientales y bases para la formulación de alternativas productivas y ocupacionales en la Región Delta. J. Adámoli y A.I. Malvárez (Eds.). Informe Técnico UBACyT N° 135. Pp. 43-60.
Willianson, M. and K. Brown . ( 1986). the analysis and modelling of british invasions. Philosophical transactions of the royal society of London, series b, 314: 505-522.
Fig. 1. Area de estudio: El Bajo Delta del Río Paraná