Estudios paleoagrostológicos comparativos en sedimentos cenozoicos argentinos
Alejandro F. ZUCOL1 , Mariana BREA1 y Esteban PASSEGGI1
1Laboratorio de Paleobotánica, CICYTTP-Diamante (CONICET), Materi y España sn, (3105), Diamante, Entre Ríos, Argentina. E-mail: cidzucol@infoaire.com.ar
Abstract: Comparative paleoagrostological studies in Argentinean Cenozoic sediments. One-thirds of the Earth's ecosystems are closely linked with grass-dominated vegetation. The evolutionary history of this ecosystem can be understood by comparative studies of the biotic component, but a paleontology approach is necessary to the major understanding of these evolutionary processes. The different methodologies applied in paleoagrostological studies are detailed in the present contribution; with special emphasis of phytolith analysis application in paleograsslands reconstruction. Paleobotanical and palynological background related to the South America Cenozoic paleoagrostology are summarized, and the different topics of these studies implementing at the present time in the Tertiary and Quaternary sediments of Argentina are described.
Key word: Paleograssland. Paleoagrostolgy. Phytolith. Cenozoic. Argentina.
Palabras clave: Paleoagrostología. Fitolitos. Cenozoico. Argentina.
Los estudios fitolíticos como herramienta para el conocimiento de los paleopastizales
Los primeros registros sobre fitolitos de sedimentos de argentina los realiza Ehrenberg (1843, 1854), quien determinó fitolitos en muestras de humus adheridas a plantas que recogiera la expedición del Beagle en Patagonia y Tierra del Fuego, como así también, en muestras de un sedimento utilizado como pigmento por los indígenas que recogiera Darwin. Entre las determinaciones que realizara Ehrenberg de material pampeano recolectado por Darwin figuran también, muestras de limos de Monte Hermoso, de limos adheridos a restos fósiles de mamíferos de Punta Alta (Bahía Blanca) y de un diente de mastodonte de las barrancas del río Carcarañá (Santa Fe), información que sería incluida por Darwin (1846) para la realización de su reporte sobre las observaciones geológicas realizadas en Sudamérica.
En Argentina los estudios fitolíticos comienzan con las investigaciones del Dr. Joaquín Frenguelli (1930), quien ya a partir de 1925 mencionara la importancia del estudio micropaleontológico de los loess y limos pampeanos; si bien su gran impulso, es a partir de los trabajos realizados por la Dra. Hetty Bertoldi de Pomar, quien entre 1969 y 1983, investigó los aspectos metodológicos de esta disciplina, como así también la presencia de fitolitos en los vegetales y en distintos tipos de depósitos sedimentarios (Bertoldi de Pomar 1970, 1971, 1972, 1974, 1975, 1976, 1980, 1983). Posteriormente, los trabajos científicos relacionados con el tema han sido escasos, y por lo general orientados a otras temáticas, como son los estudios anatómicos vegetales -en su aspecto neontológico- (Bertoldi de Pomar y Tur, 1970) o los estudios mineralógicos, pedológicos, sedimentológicos o de micropaleobotánica -en su aspecto paleontológico- (Tecchi, 1983 a y b; Andreis, 1972; Mazzoni, 1979; Peinemann y Ferreiro, 1972 a y b; Di Paola y González, 1992; Passeggi, 2000) y su aspecto metodológico (Zucol, 1992, 1995, 2001, Zucol y Brea, 2000 b).
Es a partir de la década del noventa, que se comienzan a realizar distintos tipos de estudios de los fitolitos, contemplando los aspectos de su producción y distribución en los vegetales, su incorporación al sustrato, su dispersión, conservación y presencia en distintos tipos de ambientes sedimentarios. El estudio integral de estos conocimientos es aplicado posteriormente en investigaciones paleobotánicas, paleoecológicas, pedológicas, paleolimnológicas y arqueológicas.
En el marco de estos estudios es que se han implementado las investigaciones tendientes al desarrollo de las distintas áreas de la Fitolitología y su aplicación en estudios paleoecológicos, paleoclimáticos y paleoagrostológicos.
Parte de esta tarea conforma el estudio de diferentes grupos de gramíneas actuales (Zucol, 1996 a y b; 1998; 1999; 2000; 2001), seleccionados a partir de sus abundancias relativas y sus requerimientos térmicos, que pertenecían a una región acotada: la pradera entrerriana. Estos grupos fueron procesados mediante diferentes metodologías en lo que se puede denominar una “fosilización in-vitro ” y estudiados cual si fueran restos fósiles, haciéndose hincapié en los granos de polen, los fitolitos, la anatomía foliar (en especial la disposición de los elementos epidérmicos) y la presencia de los elementos epidérmicos con elevada posibilidad de preservación ante las condiciones de fosilización. Los resultados, si se tiene en cuenta que desde un comienzo se conocía la filiación sistemática y los requerimientos mesológicos de las especies, permitió establecer el poder discriminante de los distintos caracteres estudiados y las posibilidades que estos brindan al momento de realizar inferencias paleoecológicas y paleoclimáticas (Zucol, 1996 a).
En las etapas posteriores a esta tarea, se realizó una búsqueda paleoagrostológica en sedimentos cenozoicos de la misma área que fuera estudiada neontológicamente. Estos estudios de búsqueda y prospección fueron implementados mediante análisis micropaleobotánicos de sedimentos miocenos del litoral argentino (Zucol y Brea, 2000 a y c), pliocenos, pleistocenos (Zucol y Brea, 2000 b; 2001; Brea y Zucol, 2001) y holocenos. Estos estudios guardan también relación con los análisis paleobotánicos realizados en restos fósiles petrificados e impresiones foliares con afinidad graminoide que han sido hallados en sedimentos continentales y marinos de la cuenca del río Paraná (Zucol y Brea, 1999).
La prospección realizada hizo especial hincapié en los análisis fitolíticos de perfiles sedimentarios levantados en distintos puntos de la cuenca del río Paraná en la provincia de Entre Ríos, tendientes a abarcar las distintas formaciones geológicas; complementa esta metodología un análisis paleopalinológico, de modo de obtener el marco paleoflorístico en donde se desarrollaron los representante de las familias productoras de fitolitos, tales como Poaceae, Cyperaceae, Arecaceae, Podostemaceae, entre otras con menores aportes, a modo de prospección. Esta metodología se ha implementado también en el estudio de sedimentos cenozoicos de las provincias de Buenos Aires (Osterrieth y Zucol, 2000; Zucol y Osterrieth, 2000) y del Chubut (Zucol, et al., 1999; 2001 a y b, 2002).
Esta prospección buscó abarcar tres momentos claves en la historia evolutiva de los ecosistemas dominados por gramíneas, o en forma más amplia, de los paleopastizales: su origen paleo/neógeno, su evolución e irrupción de vegetales tipo C 4 como integrantes dominantes de estos ecosistemas durante el neógeno y el desarrollo reciente de estas comunidades en vinculación directa con la actual composición y distribución florística.
Debido al estado avanzado de la prospección paleoagrostológica, es que se cuenta con la suficiente información como para comenzar una nueva etapa en estos estudios. Esta etapa, abarcaría el análisis fitolítico de sedimentos terciarios y cuaternarios, profundizando y focalizando estos estudios en sedimentos que a la luz de esta prospección presentan las mayores potencialidades para su análisis. De esta manera, se avanzaría en el estudio de la historia evolutiva de los paleopastizales de una de las regiones socio-económicamente más importantes de Argentina: la región pampeana; como así también, sobre los fenómenos paleoclimáticos y paleoecológicos que condicionaron su desarrollo en un pasado más remoto.
Antecedentes paleobotánicos y paleopalinológicos de los paleopastizales en Sudamérica
Los primeros registros palinológicos de las gramíneas en Sudamérica han sido asignados al Paleoceno (Jacobs et al., 1999); si bien es para el Eoceno donde se registra su presencia mediante estudios de fitolitos (Andreis, 1972) y para la transición Eo-Oligoceno donde la presencia de los paleopastizales se hace claramente dominante (Zucol et al., 1999) y encuentra su correlato en los cambios denticionales de los paleovertebreados herbívoros (Jacobs et al., op cit .) como consecuencia de adaptaciones coevolutivas (Stebbins, 1981). En un comienzo con una marcada preponderancia de gramíneas C 3 (Oligoceno – Mioceno medio) para posteriormente tener una composición mixta C 3 /C 4 (Mioceno medio a superior) que evidenciará un predominio de gramíneas C 4 para el Plioceno (MacFadden et al., 1996) y parte del Pleistoceno (MacFadden et al., 1999). Es durante este último lapso donde se evidencia la primera glaciación (Burnham y Graham, 1999) para continuar con los ciclos glaciales/interglaciales cuaternarios.
El conocimiento de las paleofloras durante este intervalo (Pleistoceno-Holoceno) es de suma importancia para comprender la historia evolutiva de los pastizales que actualmente se distribuyen en la región pampeana; como así también su composición y origen, ya que en estos pulsos climáticos se establecieron numerosos fenómenos de migración y mezcla de floras en respuesta a los cambios del medio ambiente. Los “depósitos pampeanos” del Pleistoceno/Holoceno abarcan este período de tiempo geológico, es por ello y por su abundante fertilidad fitolítica, que su estudio adquiere un importante valor al momento de intentar interpretar la historia evolutiva de los pastizales regionales, y en la contribución al conocimiento de los fenómenos de extinciones masivas que sucedieron durante este período con la megafauna mamífera, que en su mayoría tenían un régimen alimenticio herbívoro.
En correspondencia a los estudios realizados en la vegetación y sedimentos neógenos de la región litoral y pampeana, se estableció la búsqueda y prospección de sedimentos cenozoicos (paleógenos) de similares características. Al respecto la localidad fosilífera de Gran Barranca, situada en la barranca sur del lago Colhué Huapí (provincia del Chubut), compuesta principalmente por los sedimentos de la Formación Sarmiento, ha demostrado ser una de las localidades más aptas para este tipo de estudio. Por una parte, la presencia de fitolitos observada por Mazzoni (1979) ha sido ratificada a lo largo de toda la secuencia sedimentaria en los análisis preliminares (Zucol et al., 1999; 2001, 2002). Por otra parte, Pascual y Oderman (1971 a), como así también Andreis (1972), concluyen que durante el Eoceno Medio (Formación Muster) de Patagonia, la paleofisionomía estaba configurada por “la presencia de amplias llanuras surcadas por ríos temporarios de cauce divagante y con poca competencia de las corrientes”. Mientras que en lo referente al ambiente de depositación, Andreis et al. (1975) y Spalletti y Mazzoni (1979) postulan que “el Grupo Tobas Sarmiento representa un depósito formado esencialmente bajo condiciones subáereas, que puede ser interpretado como un paleoloess de características similares al pleistocénico y holocénico de la provincia de Buenos Aires” y que “ los fenómenos de transporte y depositación de las sedimentitas ha sido la movilización por agentes eólicos –y también ácueos- en regiones semiáridas a húmedas de bajo gradiente, que generaron sedimentos muy similares a los loess y limos que se formaron durante el Cuaternario en la región pampeana”. Spalletti (1992) plantea que “las regiones de sabana, estepa y pradera parecen ser sitios más que apropiados para la generación de los depósitos loéssicos”.
En este sentido, Behrensmeyer et al. (1992), hacen referencia a los restos hallados en diferentes tipos de sedimentos y su contexto paleoambiental y taxonómico. Con respecto a los ambientes eólicos, y en especial a los depósitos loéssicos, citan como principales microrestos vegetales a los fitolitos. Retallack (1982) encuentra que estos sedimentos se asocian al desarrollo de pastizales abiertos en el Oligo-Mioceno de Norteamérica, mientras que Wu y Goo (1985) y Kukla (1975, 1989) establecen similar relación con estepas y márgenes de desiertos del Pleistoceno de China y E de Europa. Estos autores mencionan también que en estos sedimentos loéssicos pleistocenos es común la presencia de invertebrados terrestres, estructuras caulinares, carbones y fitolitos, mientras que los palinomorfos se encuentran pobremente representados.
Esta asociación fosilífera se encuentra también presente en los sedimentos cuaternarios pampeanos. En particular, los sedimentos loéssicos de la región litoral argentina (Pampa Norte), que se caracterizan por la elevada abundancia de fitolitos y la presencia de invertebrados continentales.
Por último, la abundancia de restos de paleovertebrados (Kay et al., 1999), pertenecientes a distintas edades mamífero a lo largo de todo el perfil de la Gran Barranca, permite integrar los análisis fitolíticos a un estudio paleoecológico más abarcativo, como así también, al estudio de pautas coadaptativas entre los paleopastizales y los paleovertebrados herbívoros.
Estado actual de los análisis paleoagrostológicos
Los antecedentes previos y la tarea de investigación llevada a cabo por este grupo de investigación, abarcó:
El análisis de distintas especies de gramíneas seleccionadas por sus características mesológicas, sus distintos tipos de requerimientos térmicos y área de distribución. Estas especies fueron estudiadas en un área natural acotada y en un tiempo determinado, con la finalidad de establecer que órganos o parte de ellos poseían potencialidad de preservarse en distintos supuestos procesos de fosilización. Así como también establecer el valor diagnóstico de cada uno con respecto a la sistemática de la familia y las posibilidades de establecer inferencias paleoclimáticas y paleoecológicas a partir de los mismos (Zucol 1996 a).
La búsqueda paleoagrostológica para definir las formaciones geológicas con potencialidad de estudio, con especial hincapié en los análisis microfosilíferos. De esta manera se realizó el análisis de muestras sedimentarias de distintos sedimentos cenozoicos de Argentina, con la finalidad de establecer la fertilidad potencial de estos sedimentos en restos de naturaleza graminoide.
En la actualidad se ha arribado a las primeras conclusiones de esta tarea, que posibilitan establecer las bases para iniciar una tercera etapa, la del análisis detallado de los sedimentos seleccionados. Si bien estas tareas previas deberán continuarse con el fin de ampliar tanto los conocimientos de distribución y características diagnósticas de los fitolitos en los vegetales actuales, como así también, la potencialidad paleoagrostológica de distintos sedimentos cenozoicos de argentina. Ambas líneas de investigación, requieren ampliar sus estudios debido a que en el campo neontológico, los estudios sobre fitolitos han sido por lo general realizados en el marco de estudios anatómicos que no han contemplado a los fitolitos como formas particulares con potencialidad de definir taxa por la asociación de sus distintos componente morfológicos, por una parte; y por que, el mayor porcentaje de estudios microfosilíferos de sedimentos cenozoicos de argentina se han realizado sobre la base de análisis paleopalinológicos. Estos análisis utilizan una metodología de procesamiento de la muestra que no es la más adecuada para la concentración y conservación de restos silíceos como los fitolitos y las diatomeas, por lo cual, resulta difícil establecer la fertilidad fitolítica a partir de estudios paleopalinológicos previos, pudiendo solo establecerse si esos sedimentos poseían granos de polen de gramíneas y en que porcentajes.
Para la implementación de la nueva etapa de estudio que se esta realizando en la actualidad se plantearon una serie de premisas de trabajo, que se pueden enunciar como:
- Las características deposicionales y ambientes de sedimentación de una secuencia sedimentaria son particulares dentro de los marcos espacio – temporales de la misma o de sus componentes.
- Al conocimiento de estas características, se puede llegar mediante el análisis de una serie de factores: abióticos, que abarcan los conocimientos paleoclimáticos, del sustrato de depositación, características de depositación y factores sedimentológicos; y los bióticos, referidos a los conocimientos paleoflorístico, paleofaunísticos y paleoecológicos.
- El conjunto de los conocimientos de los factores abióticos y bióticos, permite definir un sistema deposicional característico de la secuencia sedimentaria o de sus componentes cuando la secuencia se encuentre dividida en unidades de menor magnitud.
- La comparación de secuencias sedimentarias con diferentes características espacio – temporales, debe fundamentarse en la comparación de sus respectivos sistemas deposicionales.
- Los fitolitos son considerados los principales microrestos paleobotánicos de depósitos loéssicos y ambientes de sedimentación eólica; en asociación con la presencia de restos fósiles de paleovertebrados e invertebrados terrestres, estructuras caulinares y microrestos de carbones, mientras que los palinomorfos se encuentran pobremente representados.
Estas premisas pueden resumirse en el siguiente enunciado o hipótesis principal de trabajo:
-Los análisis fitolíticos enmarcados con estudios paleontológicos de vertebrados e invertebrados, conjuntamente con las características sedimentológicas, permiten en su conjunto establecer las condiciones paleoecológicas, paleoambientales y paleoclimáticas de los sistemas de depositación de sedimentitas cenozoicas loéssicas. La definición de estos sistemas de depositación, sobre la base de sus componentes paleontológicos, características sedimentológicas y de sus condiciones paleoecológicas, paleoambientales y paleoclimáticas, permite la comparación de secuencias sedimentarias cenozoicas no coetáneas o coetáneas de diferentes localidades de estudio.
Para la evaluación de estos enunciados, se han planificado una serie de actividades que abarcan el análisis de sedimentitas loéssicas terciarias y cuaternarias, definiendo sus sistemas de depositación y su posterior comparación.
Selección de las áreas y materiales de estudio
Los estudios paleobotánicos del terciario de Sudamérica (Romero, 1986) muestran la presencia de dos tipos principales de paleofloras (una tropical o cálida y otra templado-austral) y una vegetación de mezcla distribuida en los ecotonos de ambas. A lo largo de este período la vegetación tropical con una amplia distribución durante el Terciario inferior, comienza una retracción en el lapso frío del Eoceno – Oligoceno, con el avance de la paleoflora templado fría. A partir del óptimo climático del Mioceno, y como consecuencia de los cambios orogénicos y climáticos asociados del Mio-Plioceno, se inicia un período de desertización, con la aparición de una vegetación xerófita que separa estas dos paleofloras principales en una cuña en sentido SE – NO desde la región patagónica. Estos tres componentes paleoflorísticos son los que originaron la flora actual, luego de los fenómenos de migración y mezcla ocurridos como consecuencias de los ciclos climáticos del Cuaternario.
Con la finalidad de establecer las formaciones que sedimentológica y paleontológicamente se adecuaban a estos estudios paleoagrostológicos, se implementó la búsqueda en dos campos: el primero, basado en el estudio sobre el terciario, abarcó la búsqueda de paleocomunidades dominadas por gramíneas o paleopastizales, con la finalidad de ampliar los conocimientos sobre el origen y la temprana evolución de este tipo de paleocomunidades. El segundo campo, referido al estudio sobre el cuaternario, que adquiere suma importancia debido a su estrecha vinculación con el estudio del pasado geológicamente más inmediato de una de las comunidades dominadas por gramíneas más características y de mayor importancia florística y económica de Argentina: la provincia fitogeográfica Pampeana ( sensu Cabrera y Willink, 1980).
La prospección en sedimentitas terciaria, se realizó principalmente en la provincia del Chubut (Zucol, et al., 1999; 2001 a y b, 2002), como así también en el litoral argentino (Zucol y Brea, 2000; 2001); mientras que la prospección en sedimentitas cuaternarias, abarcó las provincias de Buenos Aires y Entre Ríos (Osterrieth y Zucol, 2000; Zucol y Osterrieth, 2000; Zucol y Brea, 2000 b; 2001b; Brea y Zucol, 2001).
Para la selección del área de estudio, sobre la base de los análisis fitolíticos de sedimentos terciarios, se consideró a la localidad fosilífera de Gran Barranca la más adecuada para esta finalidad. La selección de esta localidad fosilífera, y en especial, de los sedimentos de la Formación Sarmiento, se fundamenta en la presencia de un registro fitolítico continuo a lo largo de toda la secuencia sedimentaria (Zucol et al., 1999; 2001). Sumado a los registros fitolíticos, estos depósitos son de gran importancia por sus características petrográficas, estratigráficas y paleoambientales (Andreis et al., 1975; Spalletti y Mazzoni, 1979; Mazzoni, 1979) que analogan estos sedimentos con los depósitos loéssicos pampeanos del cuaternario; como así también, por representar un lapso de tiempo geológico en donde los paleopastizales comienzan a ser de importancia en la composición paleoflorística sudamericana (Jacobs et al. 1999). Por otra parte estos sedimentos poseen una elevada riqueza paleofaunística, con la presencia de las SALMAs (South American Land Mammal Ages), Barranquense, Mustersense, “Astraponoteen plus superieur”, Deseadense y Colhuehuapense. Estas SALMAs se caracterizan por una tendencia hacia una mayor hipsodoncia a lo largo de esta secuencia sedimentaria; al respecto, esta tendencia parece haber sucedido en un rápido pulso evolutivo durante la transición Eoceno-Oligoceno, a la luz de los resultados obtenidos mediante la reciente calibración temporal de la secuencia de Gran Barranca (Kay et al., 1999, 2001), lo cual se encuentra vinculado cronológicamente con un marcado cambio climático, y la aparición de las primeras glaciaciones del Oligoceno (Zarchos et al., 2001).
Simpson (1940), precisó el intervalo de mayor transformación faunística hacia la hipsodoncia, con posterioridad al Mustersense; y vinculó la hipsodoncia (Simpson, 1953: 254) con los hábitos alimentarios de animales herbívoros, cuya dieta se compone por alimentos con elevado porcentajes de fibras y sílice. Estas observaciones fueron posteriormente ratificadas (Baker et al ., 1959; Healey y Ludwig, 1965; Barnicoat y Hall, 1960), identificándose a los agentes causantes del desgaste de las coronas dentarias de ungulados herbívoros como: abrasivos intrínsecos (fitolitos) y abrasivos extrínsecos (contaminaciones minerales de la ingesta).
El fenómeno de la transición evolutiva precoz hacia una mayor hipsodoncia en faunas patagónicas, durante el lapso de tiempo geológico desde el Casamayorense y el Deseadense fue señalado nuevamente por Pascual y Odreman (1971 a). Entre las posibles causas de este fenómeno, los autores mencionan: a los cambios en las condiciones fisionómicas, relacionadas con el levantamiento regional producido después del Mustersense (Pascual y Odreman, 1971 b), al establecimiento de una estepa herbácea (Pascual y Odreman, 1971 a), y a la depositación de sedimentos piroclásticos con abundantes clastos silíceos abrasivos.
Janis (1984) propone una metodología para el uso de conjuntos de ungulados como registro de cambios ambientales. Más tarde (Janis, 1988) establece que la hipsodoncia en ungulados guarda relación con factores de dieta y ambiente. Por su parte, Stebbins (1981) remarca la interacción entre la evolución de las gramíneas y los fenómenos de hipsodoncia en mamíferos como uno de los principales factores de su coevolución; y menciona como uno de los registros coevolutivos más antiguos, los hallados en Patagonia.
Estudios tales como el de los fitolitos en coprolitos de herbívoros (Stromberg y Stidham, 2001), así como del desgaste microscópico y los abrasivos preservados en la superficie oclusal de las coronas dentales (Fox et al ., 1996; Gobetz y Bozarth, 2001; Semprebon, 2001) pueden contribuir a determinar la importancia que han tenido los agentes abrasivos intrínsecos y extrínsecos con respecto a la hipsodoncia. No obstante, la aplicación de estos métodos para el caso de la Formación Sarmiento requiere la determinación detallada del contenido fitolítico en los distintos niveles sedimentarios.
Según Spalletti y Mazzoni (1979), la Formación Sarmiento consiste en una secuencia de sedimentitas piroclásticas de primero y segundo ciclo, de tonalidades pálidas; que constituyen estratos espesos, tabulares, y en su mayoría, macizos de homeoconglomerados, tobas y chonitas. Estos autores proponen la designación de tres unidades, que de base a techo son:
-Miembro Gran Barranca: formado principalmente por chonitas cineríticas y bentoníticas, con escasa participación de chonitas sensu stricto y tobas, y total ausencia de homeoconglomerados. Los estratos son muy potentes y macizos. Con abundantes evidencias paleoedáficas.
-Miembro Puesto Almendra: dispuesto en discordancia erosiva sobre el anterior. En su sección inferior, está constituido por homeoconglomerados, tobas y en menor proporción chonitas cineríticas, bentoníticas y sensu stricto; con presencia de una serie de evidencias paleoedáficas, como cutanes, pedotúbulos y poros. La sección superior se caracteriza por la abundancia de chonitas cineríticas y la ausencia de las tobas; intercalaciones de niveles de carbonatos (costras de tosca).
-Miembro Colhué Huapí: dispuesto en concordancia respecto al anterior. Integrado por chonitas cineríticas y tobas. En el tercio inferior se destaca la presencia de horizontes tabulares de paleosuelos.
El estudio de fitolitos en la Formación Sarmiento es una oportunidad única para conocer el temprano (Eoceno) desarrollo de ecosistemas de pastizales ocurrido en Patagonia, en comparación con otras regiones del mundo (Janis et al., 2002, Wooler, 2002).
La Formación Sarmiento es probablemente la primera unidad pre-Cenozoica tardía en ser reconocida como una loessita en razón de sus relevantes y numerosos atributos comunes con el Loess Pampeano del Cuaternario, entre ellos la abundancia y variedad de silicofitolitos (Andreis et al., 1975, Spalletti y Mazzoni, 1979, Mazzoni, 1979). La amplia gama de estudios multidisciplinarios que se están realizando en dicha unidad desde hace varios años en magnetoestratigrafía, cronología isotópica, sedimentología, icnología, paleopedología, paleontología de vertebrados, tefrocronología, etc. (Bellosi, et al., 2001, 2002 a y b; Carlini et al., 2002 a y b ; Dozo, et al., en prensa; Kay, et al., 1999, 2001 a y b) posibilitan darle un marco geológico, paleoambiental, paleobiológico y paleoecológico adecuado a los resultados de los estudios fitolíticos de estos sedimentos.
A pesar de las extendidas exposiciones de secuencias paleógenas y neógenas tempranas en Argentina, el conocimiento de las paleofloras de esa edad es poco conocido (Romero, 1986), en razón de la escasez de restos de hojas fosilizadas y de palinomorfos debido al carácter oxidante de los ambientes sedimentarios continentales. Por ello, la aproximación fitolitológica al conocimiento de las comunidades vegetales antiguas resulta una herramienta sumamente valiosa.
En lo que respecta a los análisis fitolíticos cuaternarios, los “sedimentos pampeanos” resultan ser los más adecuados para esta tarea, por su composición mineralógica como por su vinculación areal con la provincia fitogeográfica Pampeana. De los distintos sedimentos analizados en la prospección, se seleccionó la Formación Tezanos Pinto, que fuera formalmente definida por Iriondo (1980, 1987). Hasta el momento, en estos sedimentos no se han hallado restos paleobotánicos, con excepción de las asociaciones fitolíticas descriptas en la tarea de investigación preliminar. Tampoco se cuenta con resultados de estudios paleopalinológicos, como otros sedimentos cuaternarios de la región pampeana. Esta Formación resulta de importancia para estos estudios por su elevada fertilidad fitolítica, como así también, por su área de distribución.
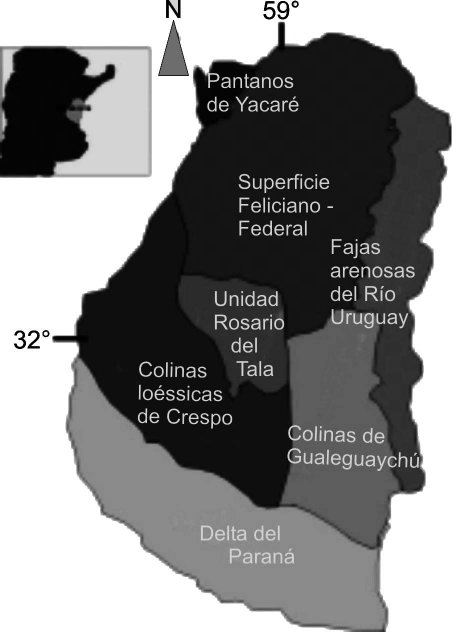
La Formación Tezanos Pinto según Iriondo (1980) constituye la unidad típica del Cuaternario de la pampa argentina. En la provincia de Entre Ríos, forma la unidad denominada “Colinas loéssicas de Crespo” (Figura 1, modificada de Iriondo, 1998). En esta área (Iriondo, 1980) esta formación cubre el relieve en forma de manto de 2 – 5 m de espesor, hacia el E ha sido parcialmente erodada y se encuentra presente sólo en las partes altas del relieve hasta cerca del río Gualeguay. Al N del arroyo Antonico (ciudad de Paraná) aparece como una capa de 1 – 1,5 m de espesor cubriendo una faja de 5 – 10 km hasta cerca de La Paz. En los ríos y arroyos de esta área este paisaje se interrumpe con el afloramiento de dos tipos de terrazas: la más antigua que corresponde a la Formación Arroyo Feliciano subyacente a los depósitos de la Formación Tezanos Pinto, y la terraza más reciente que corresponde a la Formación La Picada.
En lo que respecta a su composición, la Formación Tezanos Pinto se encuentra formada por limos eólicos (limos 71-81%; arcillas 13-29%; arenas 1-6% sensu Kröhling 1999 a, Kröhling y Orfeo 2002) depositados durante el estadío isotópico 2, en un rango de depositación que abarca entre los 36.000 a los 8.500 a A.P. (Iriondo y Kröhling, 1996). En los afloramientos del SO de Entre Ríos, la Formación Tezanos Pinto está limitada en su techo por una discordancia erosiva con la Formación San Guillermo (Iriondo, 1987) y se apoya en discordancia erosiva sobre la Formación Arroyo Feliciano (en los cauces de ríos y arroyos), la Formación Hernandarias (en el área N de las Colinas loéssicas de Crespo) o la Formación La Juanita del Grupo Punta Gorda (en el área S de las Colinas loéssicas de Crespo) (Iriondo et al., 2000). Iriondo y Manavella (1990) reconocen una serie de facies sedimentarias en esta formación en la provincia de Santa Fe: una facies de loess primario o facies eólica, una facies naranja amarillento claro con nódulos, una facies naranja mate y una facies palustre marrón amarillento grisácea. Kröhling (1999 b) diferencia dos miembros (el superior y el inferior) separados por una discordancia intraformacional visibles en perfiles potentes del centro-sur de Santa Fe. Kröhling (1998; 1999 b) mediante dataciones con termoluminiscencia define al miembro inferior con edades comprendidas entre los 36.000 y 30.000 a.A.P. (Pleistoceno tardío) y al miembro superior con edades entre los 15.000 y 8.100 a.A.P. (Pleistoceno tardío / Holoceno temprano).
En lo referente a los conocimientos paleofaunísticos de esta Formación, son escasos los restos fósiles de vertebrados principalmente en comparación de sedimentos subyacentes, (Noriega et al .,este volumen); no obstante esta unidad es portadora de abundantes restos fósiles de invertebrados continentales, con una marcada presencia de componentes malacológicos.
En este campo de estudio resulta importante contrastar las abundancias relativas, los requerimientos ecológicos y la distribución de los taxones preservados en los sedimentos cuaternarios continental y marino de la región (Aguirre, este volumen), con las características actuales del área de estudio; e integrar las inferencias basadas en la malacofauna y en los análisis isotópicos, a las obtenidas en las otras áreas de estudio.
En vinculación a ello, estos estudios se están diagramando realizando en forma coordinada con análisis de la malacofauna y otros invertebrados, incluyendo la actualización de aspectos paleoecológicos, cuantitativos, biométricos, paleobiogeográficos, geoquímicos con un enfoque paleoambiental, abarcando uno de los aspectos aún no tratados con suficiente detalle como las evidencias paleoclimáticas que los moluscos pueden brindar, especialmente durante Cuaternario tardío. Esta información resultaría con la finalidad de contrastar con otras fuentes independientes de evidencia, como las fitolíticas y sedimentológicas, así como en relación con depósitos litorales aproximadamente sincrónicos, lo que permitirá correlacionar variaciones ambientales ocurridas en los ambientes continentales estudiados. Estos estudios son además importantes para establecer si los escenarios cuaternarios a estudiar se caracterizaron por condiciones estables o por micromosaicos de cambios rápidos ambientales y climáticos, y si los moluscos de estas secuencias sedimentarias pueden ser indicadores del efecto biogeográfico de cambios climáticos.
Resultados esperados y objetivos del trabajo actual
La información paleontológica y estratigráfica es de suma importancia en los estudios sobre modelos climáticos predictivos, manejo de recursos naturales renovables y recursos florísticos; y como tal, este conocimiento biológico constituye una herramienta esencial para comprender la biodiversidad, ya que la biodiversidad en el presente sólo puede ser comprendida si se conoce su evolución histórica.
Los pastizales de América del Sur, al igual que sus contrapartes en otras latitudes, constituyen uno de los biomas terrestres más alterados, ya sea por desarrollarse en áreas potencialmente ricas para la actividad agrícola ganadera, como por ser regiones con el mayor grado de asentamientos humanos en el mundo (Soriano , 1992). En nuestro país, la Provincia Fitogeográfica Pampeana se caracteriza por la presencia de este tipo de biomas y al igual que en otras partes del mundo, sus condiciones (ecológicas, edáficas y climáticas) han posibilitado el desarrollo de distintos tipos de explotación agrícola ganadera, que ocasionaron que una gran parte de estos pastizales hayan sido reemplazados por cultivos o alterados como consecuencia de otras actividades agropecuarias (Burkart et al., 1990; León et al ., 1984).
El conocimiento de la evolución histórica de estos ecosistemas posee un doble impacto, en lo referente a la importancia de los resultados esperados, por una parte en lo que respecta a las características ecológica y botánica per se , el conocimiento de las comunidades ancestrales permite definir los componentes vegetales naturales de los pastizales, como así también los exóticos introducidos por la modificación de estos ecosistemas en forma directa o indirecta. El segundo impacto de estos conocimientos tiene vinculación con factores netamente económicos, ya que estos ecosistemas son de gran importancia desde el punto de vista agrícolo – ganadero. El conocimiento de los componen tes nativos y exóticos, como así también, su comportamiento ante las variaciones climáticas y ecológicas en su pasado histórico, son de suma importancia, al momento de la realización de acciones de manejo de estas comunidades –con distintos fines económicos- y planificación de mejoramiento de las mismas mediante introducción de especies.
Estos estudios buscan integrar y ampliar dos campos de actividades que este grupo de investigación viene desarrollando: una línea orientada al estudio fitolítico y paleobotánico en sedimentos neógenos de la región litoral (Brea, 1994, 1998, 1999; Brea y Zucol, 2001; Brea et al ., 2001; Osterrieth y Zucol, 2000; Zucol y Brea, 1999; 2000 a y b; 2001a y b; Zucol y Osterrieth, 2000), en vinculación con estudios previos orientados al estudio fitolítico de la vegetación de la región, en particular, de la pradera entrerriana (Zucol, 1996 a y b; 1998; 1999; 2000; 2001); y otra orientada al estudio en sedimentos paleógenos patagónicos (Matheos et al., 2001; Zucol, et al., 1999; 2001 a y b).
Estos estudios paleontológicos tienen un carácter interdisciplinario, tendiente al análisis de patrones de distribución de las tafofloras, correlacionando los distintos tipos de restos paleontológicos de acuerdo a las características propias de las formaciones geológicas a estudiar, y a sus características sedimentarias. Los conocimientos potenciales en el marco de estos estudios, permiten además la detección y evaluación de antiguos cambios climáticos regionales y globales y su impacto en los ecosistemas, lo cual es trascendente para el conocimiento de la actual y futura situación de nuestro planeta. Generándose además, bases de información multidisciplinaria con carácter descriptivo, que permitan la definición de modelos predictivos.
Por otra parte, conocer y posicionar cronológicamente la evolución de la vegetación tanto en el lapso Eoceno medio-Mioceno temprano en la Patagonia, como en el lapso Pleistoceno – Holoceno en la región litoral argentina, son de importancia en la interpretación plaleoflorística regional, como en el estudio de la distribución actual de los componentes florísticos.
Además, la determinación de los efectos sobre las comunidades vegetales patagónicas del cambio climático global del Eoceno tardío (“ Terminal Eocene Event ”), resulta asimismo trascendente para la evaluación de los cambios en patrones de dentición (hipsodoncia) en mamíferos.
El objetivo central del trabajo actual consiste en el análisis comparativo de las asociaciones fitolíticas y su marco paleontológico y sedimentológico, con la finalidad de establecer modelos de comparación (morfológicos, sistemáticos, ecológicos y climáticos) que permitan su aplicación al estudio comparativo de sistemas deposicionales terciarios y cuaternarios, y de esta manera se podrán reconstruir las condiciones paleoecológicas y paleoclimáticas de los diferentes sistemas.
Finalmente, estos estudios se están aplicando en dos conjuntos de actividades que abarcan los análisis de sedimentos terciarios, para lo que se seleccionó la Formación Sarmiento en la provincia del Chubut, en donde se están desarrollando además de los análisis fitolíticos, estudios de paleontología de vertebrados, icnológicos y sedimentológicos de esta formación; y las vinculadas a los análisis cuaternarios, para lo que se seleccionó las Formación Tezanos Pinto de la provincia de Entre Ríos, en donde además de los análisis fitolíticos, se complementa esta tarea con estudios de paleontología de invertebrados y sedimentológicos de esta formación.
Bibliografía
Aguirre, M. Este volumen. Moluscos del Cuaternario marino en el Sur de Entre Ríos.
Andreis, R. R., 1972. Paleosuelos de la Formación Musters (Eoceno Medio), Laguna del Mate, Prov. de Chubut. Rep. Argentina. Revista Asociación Argentina de Mineralogía, Petrología y Sedimentología, 3: 91-97.
Andreis, R. R.; Mazzoni, M. M. y L. A. Spalletti. 1975. Estudio estratigráfico y paleoambiental de las sedimentitas terciarias entre Pico Salamanca y Bahía Bustamante, Provincia de Chubut, República Argentina. Rev. Asoc. Geol. Arg. 30 (1): 85-103.
Baker, G., Jones, L. H. P. y I. D. Wardrop. 1959. Cause of wear in sheep's teeth. P, 184(4698):1583-1584.
Barnicoat, C.R. y D.M. Hall. 1960. Attrition of incisors of grazing sheep. Nature 185 (4707):179.
Behrensmeyer, A. K., Damuth, J. D., DiMichele, W. A., Potts, R., Sues, H. D. y Wing, S. L. (Eds.) 1992. Terrestrial ecosystems through time. Evolutionary Paleoecology of terrestrial plants and animals. The University of Chicago Press. Chicago & Londres. 568 p.
Bellosi, E. S., González, M. y Laza, J. 2001. Icnofaunas en paleosuelos de la Formación Sarmiento (Eoceno-Mioceno), Patagonia Central. IV° Reunión Argentina de Icnología and II° Reunión de Icnología del Mercosur. : 31. Tucumán
Bellosi, E. S., González, M., Kay, R. F. y Madden, R. H. 2002 a. El valle inciso colhuehuapense de Patagonia central (Mioceno inferior). 9ª Reunión Argentina de Sedimentología, Resúmenes, p. 49. Córdoba
Bellosi, E. S.; Miquel, S. E.; Kay, R. F. y Madden, R. H. 2002 b. Un paleosuelo mustersense con microgastrópodos terrestres (Charopidae) de la Formación Sarmiento, Eoceno de Patagonia central: significado paleoclimático. Ameghiniana 39(4):465 - 477.
Bertoldi de Pomar H. 1970. Fitolitos y zoolitos. Su significado geológico en sedimentos continentales. Boletín de la Asociación Geológica de Córdoba 1(1): 21-31.
Bertoldi de Pomar H. 1971. Ensayo de una clasificación morfológica de los silicofitolitos. Ameghiniana P, 8 (3-4): 317-328.
Bertoldi de Pomar H. 1972. Ópalo organógeno en sedimentos superficiales de la llanura santafecina. Ameghiniana 9(3): 265-279.
Bertoldi de Pomar H. 1974. Silicobiolitos en sedimentos de cauce fluviales correntinos. I Cong. Arg. de Paleont. y Bioestrat., Actas :633-639.
Bertoldi de Pomar H. 1975. Los silicofitolitos: Sinopsis de su conocimiento. Darwiniana 19(2-4): 173-206.
Bertoldi de Pomar H. 1976. Métodos de preparación de sedimentos clásticos para su estudio microscópico. I. Tratamientos previos. Revista Asociación Ciencias Naturales del Litoral 7: 1-55.
Bertoldi de Pomar H. 1980.Análisis comparativo de silicobiolitos de diversos sedimentos continentales argentinos. Revista Asociación Geológica Argentina 35(4): 547-557.
Bertoldi de Pomar H. 1983. Silicobiolitos en sedimentos del cauce del Río Paraguay. Rev. Asoc. Cs. Nat. del Litoral 14(1): 53-62.
Bertoldi de Pomar H. y N.M. Tur. 1970. Células silicificadas en gramíneas acuáticas. Revista de la Asociación de Ciencias Naturales del Litoral 1: 17-18.
Brea, M. 1994. The firts permineralised wood of Combretaceae in the Salto Chico Formation from Entre Ríos Province, Argentina. IAWA, Journal 15 (3): 328.
Brea, M., 1998. Ulminium mucilaginosum n.sp. y Ulminum artabeae n.sp., Dos leños de Lauraceae en sedimentos de la Formación El palmar, provincia de Entre Ríos, Argentina. Ameghiniana 35 (2): 193-204. Buenos Aires.
Brea, M., 1999. Leños fósiles de Anacardiaceae y Mimosaceae de la Formación El palmar (Pleistocenos superior) del departamento Concordia, provincia de Entre Ríos. Ameghiniana 36 (1): 63-69. Buenos Aires.
Brea, M. y A. F. Zucol. 2000. Lignofloras del Cenozoico Superior del Noreste Argentino. En: Aceñolaza, G. y R. Herbst (eds.), “ El Neógeno de Argentina.” Serie Correlación Geológica, 14: 245 – 253.
Brea, M. y A. F. Zucol. 2001. Estudio paleobotánico del Parque Nacional El Palmar (Entre Ríos, Argentina): Silicobiolitos en sedimentos continentales del Pleistoceno Superior . Ameghiniana P, 38 (4-suplemento): 29R. Buenos Aires.
Brea, M.; Zucol, A. F. y A. Scopel. 2001. Estudios paleobotánicos del Parque Nacional El Palmar (Argentina). I. Presencia de cuerpos de sílice en leños fósiles de Mirtaceae. Natura Neotropicalis , P, 32 (1): 33 - 40.
Burkart, S. E., León, R. J. C. y C. P. Movia. 1990. Inventario fitosociológico del pastizal de la Depresión del Salado (provincia de Buenos Aires) en un área representativa de sus principales ambientes. Darwiniana 30(1-4): 27 – 69.
Burnham, R. J. y A. Graham. 1999. The history of Neotropical vegetation: New developments and status. Annals of Missouri Botanical Garden , P, 86: 546-589.
Cabrera, A. L. y A. Willink. 1980. Biogeografía de América latina. Monografía de la Secretaria General de la Organización de los Estados Americanos. Serie biología Nº 13. Washington, D.C. 123 p.
Carlini, A. A., Scillato-Yané, G. J., Madden, R. H., Soibelzon, E. y Ciancio, M. 2002 a. Los Dasypodidae (Mammalia, Xenarthra) del Eoceno. 2) El conjunto de especies del Casamayorense de Cañadón Vaca, SE de Chubut (Argentina) y su relación con los que le suceden. 1º Congreso Latinoamericano de Paleontología de Vertebrados , Resúmenes: 24. Santiago, Chile.
Carlini, A. A., Scillato-Yané, G. J., Madden, R. H., Ciancio, M. y Soibelzon, E. 2002 b.Los Dasypodidae (Mammalia, Xenarthra) del Eoceno. 3) Las especies del Casamayorense de la Barranca Sur del lago Colhué Huapi, S de Chubut (Argentina): el establecimiento de los Euphractinae. 1º Congreso Latinoamericano de Paleontología de Vertebrados , Resúmenes: 24-25. Santiago, Chile.
Darwin, P., 1846. Geological observations on South America, Being the third part of the Geology of the Voyage the Beagle, during the years 1832 to 1836 . Smith elder and Co., i-vii, London. 279 pp.
Di Paola, E. C. y M. González. 1992. Silicofitolitos en secuencias continentales terciario-pleistocénicas de la provincia de San Luis. Revista de la Asociación Geológica Argentina 47(1): 111 – 112.
Dozo, M. T., Vucetich, M. G. y Candela, A. M. En prensa. Skull anatomy and neuromorphology of Hypsosteiromys axiculus , a Colhuehuapian Erethizontidae rodent from Chubut, Argentina, Journal of Vertebrate Paleontology.
Ehrenberg., C. G. 1843. Verbreitung und einfluss des mikroskopischen lebens in Süd- und Nord-Amerika. Abhandl. Berlin Akad.
Ehrenberg, C. G. 1854. Uber ein aus feinstein Kieselmehl von Infusorien bestehende Schminke der Feuerländer. Akad. D. Wissensch. zu Berlin :11-12.
Fox, C.L., Juan, J. y R.M. Albert. 1996. Phytolith analysis on dental calculus, enamel surface, and burial soil: Information about diet and paleoenvironment. American Journal of Physical Anthropology 101:101-113.
Frenguelli, J. 1925. Loess y limos pampeanos. Anales de la Sociedad Argentina de Estudios Geográficos GAEA 1: 1-88.
Frenguelli, J. 1930. Partículas de sílice organizada en el loess y en los limos pampeanos. Células silíceas de Gramíneas. Anales de la Soc. Cient. de Santa Fé 2 :1-47.
Gobetz, K. E. y S. R. Bozarth. 2001. Implications for late Pleistocene Mastodon diet from opal phytoliths in tooth calculus. Quaternary Research, P, 55:115-122.
Healey, W. B. y T. G. Ludwig. 1965. Ingestion of soil by sheep in New Zealand in relation to wear of teeth. Nature , P, 208:806-807.
Iriondo, M. 1980. El cuaternario de Entre Ríos. Revista de la Asociación de Ciencias Naturales del Litoral , 11: 125-141.
Iriondo, M. 1987. Geomorfología y Cuaternario de la provincia de Santa Fe (Argentina). D'Orbignyana 4: 1-54.
Iriondo, M.1998. Loess in Argentina: temperate and tropical. Guide N° 3 Province of Entre Ríos. International Joint Field Meeting, INQUA, 12 p. Paraná.
Iriondo, M. y D.M. Kröhling. 1996. Los sedimentos eólicos del Noreste de la llanura pampeana (Cuaternario Superior). XIII Congreso Geológico Argentino y III Congreso de explotación de hidrocarburos, Actas IV: 27-48.
Iriondo, M. y C. Manavella. 1990. Facies sedimentarias de la Fm Tezanos Pinto en el centro de la provincia de Santa Fe. En: Zárate, M.(ed.) International Symposium on Loess, properties, chronology and paleoclimatic significance of loess. INQUA, Expanded abstracts: 74-77. Mar del Plata.
Iriondo, M., Kröhling, D. y Bidegain, J., 2000. The Quaternary of SW Entre Rios, Argentina. Abstract (CD) 31th. International Geological Congress. Río de Janeiro.
Jacobs, B.F., Kingston, J.D. y L.L. Jacobs. 1999. The origin of grass-dominated ecosystems. Annals of Missouri Botanical Garden , 86:590-643.
Janis, C.M. 1984. The use of fossil ungulate communities as indicators of climate and environment. En: Brenchley P. (Ed.), Fossils and Climate . John Wiley & Sons, Ltd., London.
Janis, C.M. 1988. An estimation of tooth volume and hypsodonty indices in ungulate mammals, and the correlation of these factors with dietary preference. En: Russell, D.E.; Santoro, J.-P. y D. Sigogneau-Russell (Eds.), Teeth Revisited. Mem. Mus. Natn. Hist. Nat., Paris, (serie C), 53:367-387.
Janis, C.M., Damuth, J. y J. Theodor. 2002. The origins and evolution of the North American grassland biome: the history from the hoofed mammals. Palaeogeog. Palaeoclim. Palaeoecol., 177: 183-198.
Kay , R.F. Madden, R.H., Vucetich, M.G., Carlini, A.A., Mazzoni, M.M., Re, G.H., Heizler M. y H. Sandeman. 1999. Revised geochronology of the Casamayoran South American Land Mammal Age: Climatic and biotic implications. Proc. Nat. Acad. Of Science 96 (23): 13235-13240.
Kay, R.F., Madden, R.H., Bellosi, E., Carlini, A.A., Heizler, M., Re, G., Vilas, F. y M.G. Vucetich. 2001 a. Puesto Almendra-Colhué Huapi contact at Gran Barranca, Sarmiento Formation, Chubut, Argentina. Ameghiniana 38(4-Suplemento):35R.
Kay, R. F., Madden, R. H., Carlini, A. A., Bellosi, E., Heizler, M., Re, G., Vilas, F. y Vucetich, M. G. 2001 b. The Mustersan interval at Gran Barranca. Ameghiniana 38(4-Suplemento): 34R.
Kröhling, D.M. 1998. Loess in Argentina: temperate and tropical. Guide N° 2 North Pampa (Carcarañá river basin, Santa Fe province). International Joint Field Meeting, INQUA, 33 p. Paraná.
Kröhling, D.M. 1999 a. Upper Quaternary geology of the lower Carcarañá Basin, North Pampa, Argentina. Quaternary International 57/58: 135-148.
Kröhling, D., 1999b. Sedimentary maps of loessic units outcropping in North Pampa, Argentina. Quaternary International, 62: 49-55.
Kröhling, D. y O. Orfeo 2002. Sedimentología de unidades loéssicas (Pleistoceno Tardio-Holoceno) del centro-sur de Santa Fe. Revista de la Asociación Argentina de Sedimentología (AAS). 9 (2): 135 – 154.
Kukla, G. 1975. Loess stratigraphy of Central Europe. En: Butzer, K. W. y G. Ll. Isaac (eds.) “ After the Australopithecines.” : 99 – 188. The Hague. Mouton.
Kukla, G. 1989. Loess stratigraphy in Central China. Palaeogeog. Palaeoclim. Palaeoecol ., 72: 203 – 225.
León, R. J. C., Rusch, G. M. y M. Oesterheld. 1984. Pastizales pampeanos – Impacto agropecuario. Phytocoenologia 12(2-3): 201 – 218.
MacFadden, B.J., Cerling, T.F. y J. Prado. 1996. Cenozoic terrestrial ecosystem evolution in Argentina: Evidence from carbon isotopes of fossil mammal teeth. Palaios 11: 319-327.
MacFadden, B.J., Cerling, T.F. y J. Prado. 1999. Ancient latitudinal gradients of C 3 /C 4 grasses interpreted from stable isotopes of New World Pleistocene horse ( Equus ) teeth. Global Ecology and Biogeography 8: 137-149.
Matheos, S., Brea, M., Ganuza D. y A. Zamuner. 2001. Sedimentología y paleoecología del Terciario inferior en el sur de la provincia del Chubut, República Argentina. Asociación Argentina de Sedimentología, Revista 8 (1): 93-104.
Mazzoni, M. M. 1979. Contribución al conocimiento petrográfico de la Formación Sarmiento, barranca S del lago Colgué Huapí, provincia del Chubut. Revista Asociación Argentina de Mineralogía, Petrología y Sedimentología 10 (3-4): 33-53.
Noriega, J. I., Carlini, A. A. y E. P. Tonni. este volumen. Vertebrados del Pleistoceno tardío de la cuenca del Arroyo Ensenada (Departamento Diamante, Provincia de Entre Ríos, Argentina.
Osterrieth, M.L. y A.F. Zucol. 2000. Paleoenvirnmental reconstruction in late quaternary paleosoils of the pampean plains (Argentina). 3 rd International Meeting on Phytolith Research. Abstracts :15-16.
Pascual, R. y O. Odreman Rivas. 1971 a. Evolución de las comunidades de los vertebrados del Terciario argentino; los aspectos paleozoogeográficos y paleoclimáticos relacionados. Ameghiniana 7(3-4): 372-412.
Pascual, R. y O. Odreman Rivas. 1971 b. Las unidades estratigráficas del Terciario portadoras de mamíferos; su distribución y sus relaciones con los acontecimientos diastróficos. 5 Congreso Geológico Argentino, Actas : 293-335.
Passeggi, E. 2000. Caracterización Sedimentológica del Material Parental de los Suelos Asociados a los Depósitos de Cauce del Tramo Medio del Río Paraná . Tesis doctoral. Universidad Católica de Santa Fe. 216 p.
Peinemann, N. y E. A. Ferreiro. 1972 a. Fitolitos en suelos pampeanos. Influencia sobre las propiedades físicas determinantes de la evolución de la estructura. Ciencia del suelo 8(2): 135 - 141.
Peinemann, N. y E. A. Ferreiro. 1972 b. Accumulation of amorphous silica in the Pampas plains. Zeitschrift für Pflanzenemährung und Bodenkurde 132: 1 – 4.
Retallack, G. J. 1982. Paleopedological perspectives on the development of grasslands during the Tertiary. Proceeding of the 3th North American Paleontological Convention 2: 417 – 421.
Romero, E.J. 1986. Paleogene phytogeography and climatology of South America. Annals of the Missouri Botanical Garden 73: 449-461.
Semprebon, G. 2001. A new method of reconstructing ungulate dietary behavior with applications to fossil equids. Journal of Vertebrate Paleontology 21(3):100A.
Simpson, G. G. 1940. Review of the mammal-bearing Tertiary of South America. Proceeding of the American Philosophical Society 83(5):649-709.
Simpson, G. G. 1953. The Major Features of Evolution. Columbia University Press, N.Y.
Soriano, A. 1992. Río de la Plata grasslands. En: Coupland, R. T. (ed.) “ Natural grasslands: Introduction and western hemisphere. Ecosystem of the world .” : 367 – 407. Elsevier.
Spalletti, L. A. 1992. El loess y el problema de la identificación de las loessitas. Revista del Museo de La Plata (nueva serie) –secc. Geología n° 102- 45-56.
Spalletti, L. A. y M. M. Mazzoni. 1979. Estratigrafía de la Formación Sarmiento en la barranca sur del Lago Colhué Huapí, provincia del Chubut . Revista Asociación Geológica Argentina 34(4): 271-281.
Stebbins, L. G. 1981. Coevolution of grasses and herbivores. Annals of Missouri Botanical Garden 68: 75-86.
Stromberg, C. A .E. y T. A. Stidham. 2001. Dung beetle brood balls and notoungulate diet. Journal of Vertebrate Paleontology 21(3):105A.
Tecchi, R. A.1983 a. Contenido de silicofitolitos en suelos del sector sudoriental de la Pampa Ondulada. Ciencia del suelo 1(1): 75 - 82.
Tecchi, R. A.1983 b. Distribución de silicofitolitos en dos perfiles de suelo de la Pampa Ondulada. Ciencia del suelo 1(2): 93 - 95.
Wooler, M.J. 2002. Introduction to the reconstruction and modeling of grass-dominanted ecosystems. Palaeogeog. Palaeoclim. Palaeoecol. , 177: 1-3.
Wu, Z. y F. Gao. 1985. The formation of loess in China. En: Liu, T. (ed.). “ Quaternary Geology and environment of China.” : 137 – 138. Beijing, China Ocean Press.
Zarchos, J., Pagani, M., Sloan, L., Thomas, E. y K. Billups. 2001. Trends, rhytms, and aberrations in global climate 65 Ma to present. Science 292: 686-693.
Zucol, A. F. 1992. Microfitolitos: I. Antecedentes y terminología. Ameghiniana 29(4): 353-362.
Zucol, A. F. 1995. Microfitolitos: II. Análisis de las clasificaciones. Ameghiniana 32(3): 243-248.
Zucol, A. F. 1996 a. Estudios morfológicos comparativos de especies de los géneros Stipa, Panicum y Paspalum (Poaceae), de la Provincia de Entre Ríos. Tesis doctoral, U.N.L.P. 558 pp.
Zucol, A. F. 1996 b. Microfitolitos de las Poaceae argentinas: I. Microfitolitos foliares de algunas especies del género Stipa (Stipeae: Arundinoideae: Poaceae), en la provincia de Entre Ríos. Darwiniana 34 (1-4): 151-172.
Zucol, A. F. 1998. Microfitolitos de la Poaceae argentinas: II. Microfitolitos foliares de algunas especies del género Panicum (Paniceae: Panicoideae: Poaceae), en la provincia de Entre Ríos. Darwiniana 36 (1-4): 29-50.
Zucol, A. F. 1999. Fitolitos de las Poaceae argentinas. IV. Asociación fitolítica de Cortaderia selloana (Danthonieae: Arundinoideae), de la provincia de Entre Ríos (Argentina). Natura Neotropicalis 30 (1-2): 25-33.
Zucol, A. F. 2000. Fitolitos de Poaceae argentinas: III. Fitolitos foliares de algunas especies del género Paspalum (Paniceae: Panicoideae: Poaceae), en la provincia de Entre Ríos. Darwiniana 38 (1-2): 11-32.
Zucol, A. F. 2001 a. Fitolitos: III. Asociaciones fitolíticas de Piptochaetium montevidense (Stipeae: Arundinoideae: Poaceae). Una nueva metodología descriptiva. Boletín de la Sociedad Argentina de Botánica 36 (1-2): 69 – 85.
Zucol A. F. y M. Brea. 1999. Una Bambusoideae petrificada de la Formación Ituzaingó (Plioceno), Paraná, Entre Ríos. Reunión anual de comunicaciones Científicas de la Asociación Paleontológica Argentina. Tucumán.
Zucol, A. F. y M. Brea. 2000 a. Análisis fitolítico de la Formación Paraná (Mioceno superior) en el Departamento Diamante, Entre Ríos, Argentina. II Congreso Sedimentológico Latinoamericano y VIII Reunión Argentina de Sedimentología, Resúmenes: 190-191. Mar del Plata.
Zucol, A. F y M. Brea. 2000 b. Sistemática de fitolitos, pautas para un sistema clasificatorio. Un caso en estudio: asociación fitolítica de la Formación Alvear en su área tipo. XI Simposio Argentino de Paleobotánica y Palinología, Resúmenes: 120. Tucumán.
Zucol, A. F. y M. Brea. 2000 c. Análisis fitolítico de la Formación Paraná en la provincia de Entre Ríos. En: Aceñolaza, G. y R. Herbst (eds.), El Neógeno de Argentina. Serie Correlación Geológica, 14: 67 – 76.
Zucol, A. F. y M. Brea. 2001. Asociación fitolítica de la formación Alvear (Pleistoceno Inferior), Entre Ríos, Argentina. Ameghiniana 38 (4-suplemento): 49R. Buenos Aires.
Zucol, A. F. y M. Osterrieth. 2000. Análisis fitolítico del Holoceno tardío de Mar Chiquita, Buenos Aires, Argentina. II Congreso Sedimentológico Latinoamericano y VIII Reunión Argentina de Sedimentología. Resúmenes: 191-192. Mar del Plata.
Zucol, A. F., M. Brea y R. H. Madden. 2002. Preliminary phytolith analysis of mmz profile in the Gran Barranca sedimentary sequence (Chubut, Argentina). 4 th International Meeting of Phytolith Research, Cambridge : 34.
Zucol, A. F., Brea, M. y S. Matheos. 2001. Estudio perliminar de microrestos silíceos del Paleoceno de la Formación Salamanca (Paleoceno) en la provincia del Chubut, Argentina. Ameghiniana 38 (4-suplemento): 49 – 50R. Buenos Aires.
Zucol, A. F., Mazzoni, M. M. y R. H. Madden. 1999. Análisis fitolíticos en la secuencia sedimentaria de Gran Barranca, Chubut. Ameghiniana 36 (4-suplemento): 43R. Buenos Aires.
Zucol, A. F., Brea, M., Carlini, A. A. y R. H. Madden. 2001. Análisis fitolíticos en la secuencia sedimentaria de Gran Barranca, Chubut, Argentina: II. El Miembro Colhué Huapi (Formación Sarmiento). Ameghiniana 38 (4-suplemento): 50R. Buenos Aires.
Fig. 1. Mapa Geomorfológico de Entre Ríos (sensu Iriondo, 1998).